大和陽一
ア. 研究目的
これまで沖縄地方ではイチゴは栽培されていなかったが、単価の高い品目であり、高収益が期待される。本研究では沖縄地方に導入する高収益野菜としてイチゴを取り上げ、亜熱帯の気象条件を考慮した短期栽培技術を開発する。
イ. 研究方法
1. 定植時の花芽分化段階の検討(1998-1999)
1998年に‘とよのか’および‘さちのか’を野菜茶試久留米支場(現野菜花き研究部)で育苗した。8月28日以降は16時間日長、夜温摂氏20度以上で管理し、花芽の自然分化を抑えた。花芽分化誘導のための短日夜冷処理(自然光型人工気象室、昼/夜温摂氏30~35/13度、8時間日長)の期間を変えることにより、試験区を設定した((1)a区(短日夜冷処理なし、頂果房の花芽分化期)、(2)b区(頂果房が分化、がく片形成期)および(1)c区(頂果房と腋果房が分化))。11月5日に沖縄県農試園芸支場の鉄骨ハウス(間口8m×長さ27m)に定植した。土壌改良のため堆肥4.5t/10aとリン酸10kg/10a,元肥としてN,P,Kをそれぞれ10kg/10a施用した。栽植様式は畝幅120cm,株間25cm,条間25cm,2条千鳥植えとし、地温上昇抑制のためシルバーマルチを用いた。なお、沖縄にミカンキイロアザミウマを持ち込まないように徹底した処置を施した。
2. 定植時期の検討(1998-1999)
1998年に久留米支場で育苗した‘とよのか’と‘さちのか’を園芸支場に輸送した。定植時期の違いにより試験区を設定し((2)b~(2)f)、それぞれの定植時期に合わせて花芽分化させるために短日夜冷処理を行った。
3. 花芽分化時期と温度の関係(1999)
1999年6月1日および9月1日に園芸支場でポット受けした‘さちのか’と‘とよのか’の苗を、それぞれ6月15日および9月15日に園芸支場の圃場に仮植した。施肥量は、N,P,K,それぞれ5kg/10aとした。9月25日から11月25日の間に10日間隔で花芽分化を調査し、0:未分化、1:肥厚初期、2:肥厚中期、3:肥厚後期、4:2分期、5:がく片形成期、6:雄ずい形成期、7:雌ずい形成期、8:出蕾前、および9:出蕾後に分類した。
4. 定植時の花芽分化段階および定植後の遮光の効果(1999-2000)
久留米支場で‘とよのか’と‘さちのか’を1999年8月中旬に採苗し、ガラス温室で10.5cmポットを用いて育苗した。9月以降は、自然状態での花芽分化を抑制するために、16時間日長、夜温摂氏20度以上で管理した。花芽分化を誘導するために、短日夜冷(8時間日長、夜温摂氏13度)20(‘とよのか’では15日間)および40日間処理、ならびに低温暗黒(摂氏10度)20日間処理(‘さちのか’のみ)を行った。さらに、‘さちのか’では短日夜冷20日処理した苗に定植後トンネルにより50%遮光を行う区を設けた。11月1日に園芸支場の鉄骨ハウスに定植した。
5. 花芽分化抑制処理の効果(1999-2000)
久留米支場で‘さちのか’を1999年8月中旬に採苗し、ガラス温室で10.5cmポットを用いて育苗した。花芽分化抑制処理として、電照による16時間日長の長日処理、暖房による夜温摂氏20度の高温処理、ならびに高温長日処理を行った。さらに、花芽分化抑制処理を行わない苗の頂花房を定植後(12月7日)除去する区を設けた。9月以降、高温区と高温長日区以外は雨よけハウスで育苗した。11月1日に園芸支場の鉄骨ハウスに定植した。
6. 最適な採苗時期と低温暗黒処理の効果(1999-2000)
沖縄県農試園芸支場で‘さちのか’を1999年6月15日、7月15日、8月15日および9月15日に採苗し、10.5cmポットで育苗した。低温暗黒処理は、10月20日から摂氏13度に設定した冷蔵庫を用いて行った。11月1日に定植した。
7. 沖縄におけるポット苗の頂花房分化時期(2000)
‘さちのか’と‘とよのか’の沖縄での生産株から8月下旬に採苗し、9月8~10日に10.5cmポットに鉢上げした。育苗期は、遮光率45%の遮光資材で被覆した。処理区として、低N区(育苗中にN成分140mg/ポット施用)、高N区(同210mg/ポット)および低N・低温暗黒処理区(低N,摂氏10度の冷蔵庫で10日間処理)を設けた。
8. 沖縄に適した短期栽培における作型の検討(2000-2001)
‘さちのか’と‘とよのか’の沖縄での生産株から2000年8月25日に採苗し、10.5cmポットに鉢上げした。低温暗黒処理を行い、頂花房の花芽分化を検鏡により調査した。また、T1(10月16日定植、低温暗黒処理なし)、T1c(10月16日定植、低温暗黒処理あり)、T2(10月25日定植、低温暗黒処理なし)、T2c(10月25日定植、低温暗黒処理あり)、Tn(10月31日定植、低温暗黒処理なし)、Tn2(10月31日定植、低温暗黒処理なし、2芽仕立て)およびT1cb(10月16日定植、低温暗黒処理あり、果実遮光)区を設けた。
9. 沖縄でのイチゴ栽培に適した花芽分化処理法の検討(2001-2002)
園芸支場で2001年8月14日に‘さちのか’を採苗した。9月3~4日に10.5cmポットに鉢上げし、園芸支場の45%遮光した鉄骨ハウス内で育苗した。鉢上げ後のN施用量(低N;140mg/ポット、高N;210mg/ポット)と低温暗黒処理期間(10,15および20日間)を組み合わせて試験区を設定した。低温暗黒処理は11月1日に終了するように処理を開始した。また、沖縄に適した花芽分化処理法について検討するために、親株の前歴(前年度の生産株、11月下旬~2月中旬に摂氏4度で冷蔵して休眠打破した株)と花芽分化処理法(無処理、短日夜冷(8時間日長、夜温摂氏15度)20および36日間処理、低温暗黒(摂氏10度)15日間処理ならびに遮光(寒冷紗でさらに50%遮光)処理を組み合わせて、試験区を設定した。10月16日に処理を終了し、園芸支場の鉄骨ハウスに定植した。短日夜冷36日区では7月27日に挿し苗し、8月8日に鉢上げした。2002年に同様の試験を行った。短日夜冷処理期間は40および20日とした。
10. セル育苗の可能性と定植後の水分管理法の検討(2001)
‘さちのか’を用いて、育苗方法(50穴セルトレイ、7.5cmおよび10.5cmポットで育苗)と定植後の灌水処理(多、通常(これまでの試験と同様に週2~3回灌水)および少灌水;畝に設置したテンショメータでpF値をモニタしながら灌水)を組み合わせて試験区を設定した。10.5cmポットで育苗した区では生産株と休眠打破株から採苗し、他はすべて生産株から採苗した。園芸支場で2段階採苗により8月14日に採苗し、9月3~4日に鉢上げした(セルトレイ区はそのまま)。園芸支場の45%遮光したハウスで育苗し、10月31日に鉄骨ハウスに定植した。
11. 定植後の水分・施肥管理法の検討(2002)
園芸支場で‘さちのか’を2002年9月3~5日に採苗し、10.5cmポットを用いて育苗した。10月30日に園芸支場内の鉄骨ハウスに定植した。施肥条件(N,P,K,それぞれ10kg/10aが標準、30%減肥、2月10日以降0.5kgN/週/10aとなるよう追肥)と灌水処理(少灌水;週1回灌水、通常灌水;週3回灌水、多灌水;毎日灌水)を組み合わせて試験区を設定した。
ウ. 結果および考察
1 .定植時の花芽分化段階の検討
実験期間中暖房は行わず、ハウスのサイドは夜間も常時開放して、雨天時および強風時以外閉めなかった。また、電照は行わず、ジベレリンも使用しなかった。ハウス外の日射量(データ略)は、12月中旬~1月下旬頃に少なくなったが、2月中旬以降に多く、全体的にほぼ平年並みであると考えられた。久留米での日射量と比較すると、時期的な差はあるが、全体的には大きな差はなかった。ハウス内の気温は1月下旬~2月上旬が最も低温で、半旬別平均値で日最低気温が約摂氏12度、日最高気温が約摂氏19度であった。久留米でのハウス内気温の値と比較すると、沖縄のハウスは軒高でサイドを常に高くまで開放してあることから、日最高気温は久留米のビニルハウスよりやや低く推移した。日最低気温については、久留米では暖房しているが、久留米の方が摂氏5~10度低く推移した。
頂花房の出蕾・開花日(第1表)は、(1)a,(1)b区で12月6~7日であり、(1)c区で1週間程度早くなった。頂花房の収穫開始日は、(1)a,(1)b区より約10日早くなった。(1)c区の第1次腋花房の開花は(1)b区より2~3日早くなり、収穫は5~7日早くなった。成熟日数は、頂花房では‘とよのか’で25~27日、‘さちのか’では27~30日であった。第1次腋果房では、頂花房よりやや長くなった。久留米では沖縄より生育が遅く、頂花房では開花が9~16日遅れ、収穫開始は28~30日遅れた(データ略)。果実の成熟日数も沖縄より長期間を要し、頂花房は‘とよのか’で46~48日、‘さちのか’で44~49日であった。
収穫果実の果重・個数の積算値(データ略)は、‘とよのか’では1月下旬以降(1)b区で最高であった。1月下旬~2月末までは、次いで(1)a,(1)c区の順であったが、それ以降逆転した。‘さちのか’では3月上旬までは(1)c区で最も高かったが、それ以降は(1)b区で最も高くなった。株当たりの収量は2月末までは両品種とも約150~200g,3月末までは‘とよのか’で約300~370g,‘さちのか’で約350~420gであった。
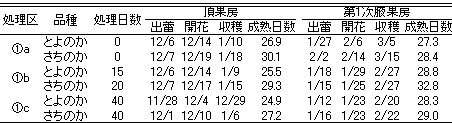
第1表 各処理区の頂果房および第1次腋果房の出蕾・開花日と頂花の成熟日数
2. 定植時期の検討
いずれの区においても定植後の苗の生育は順調であった。定植直後の出葉速度は久留米で定植した場合と同様であり、いずれの定植時期でも苗の活着については問題がないものと考えられた。
頂花房の出蕾・開花および収穫開始(第2表)は定植日が早いほど早くなり、(2)f区を除いて‘さちのか’より‘とよのか’で早かった。第1次腋花房の出蕾は(2)b区で最も早かった。最も遅れたのは、‘とよのか’では(2)f区、‘さちのか’では(2)e区であった。定植日の遅い区では、第2次腋花房の未出蕾株が多かった。また、定植日の違いによる第2次腋花房の収穫開始日の差は小さかった。
出蕾・開花および収穫開始日は、沖縄より久留米で遅くなった。また、最低気温が高くなると第1次腋花房出蕾までに多くの日数を要する関係が見られた(第1図)。開花から収穫開始までの日数(果実成熟日数)とその期間の日最低気温の平均値との関係(第2図)を見ると、日最低気温が高いと果実の成熟日数は短くなり、その関係は久留米と沖縄で同一であった。

第2表 各区の定植日と定植時の花芽分化状態ならびに頂果房および腋果房の出蕾・開花日
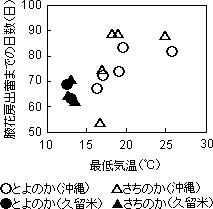
?第1図 定植から腋花房出蕾までの
日数と最低気温の関係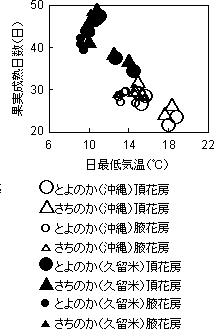
第2図 果実成熟日数と
日最低気温との関係
早く定植した区では、第1次腋花房の収穫開始までの日数がかかったため、1月下旬~2月上旬までの収量は少なく、特に(2)b区の‘さちのか’では収穫果実がほとんどなかった(データ略)。3月末までの株当たりの収穫果重は(2)c区で多く、‘とよのか’で442g/株(295kg/a)、‘さちのか’で518g/株(345kg/a)であった。
果実の硬度、糖度および酸度には時期的な変化のパターンが見られた(データ略)が、各区とも同様に変化しており、気象条件が影響するものと考えられた。‘とよのか’の果実硬度は約300g/3mmφであったが、1月下旬~2月中旬頃に120~160g/3mmφ程度になった。‘さちのか’ではさらに果実硬度が高く、収穫始めから150g/3mmφ以上で推移したが、3月に入ると100g/3mmφ程度まで低下した。果実糖度(Brix)は、‘とよのか’では収穫開始~2月中旬までは10度前後であった。‘さちのか’では2月下旬頃までは10~12度であり、‘とよのか’より高く推移した。3月上~中旬には両品種ともに8~9度程度まで低下し、その後やや上昇する傾向が見られた。果実酸度は、全期間を通じて‘とよのか’より‘さちのか’の方が低く推移した。総合的な果実品質は‘とよのか’より‘さちのか’の方が優れると考えられた。
以上の結果より、定植時期が早い方が収量は高くなり、両品種ともに沖縄では充分な収量が得られること、冬季には品質の良い商品性の高い果実が得られることが確認された。全期間を通じて、‘とよのか’より‘さちのか’の方が糖度、酸度および等の果実特性が優れ、最終的な収量も高くなる傾向が見られた。第1次腋花房の出蕾特性や果実糖度、酸度の推移を考慮すると、両品種とも沖縄での栽培に適した定植時期として11月1日前後が1つの目安となると考えられた。また、以上の結果は、久留米で育成し花芽分化させた苗について得られたものであり、沖縄で低温遭遇が不十分な親株から得られた苗についてはさらに検討が必要であると考えられた。
3. 花芽分化時期と温度の関係
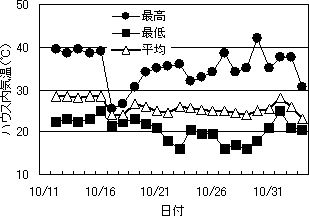
第3図 ハウス内気温の推移
10月15日以降、ハウス内の気温は徐々に低下した(第3図)。
頂花房の花芽分化は、両品種ともに採苗時期に関係なく10月5日以降確認された(データ省略)。しかし、個体間差が大きく、差は10月25日以降になって小さくなった。第1次腋花房の花芽分化は、両試験区で両品種ともに11月はじめ頃から確認され、頂花房に比べ個体間差は小さかった。
以上のことから、花芽分化は採苗時期や生育にはあまり影響されず、温度による影響が大きいと考えられた。沖縄における頂花房の分化時期は10日20日前後、第1次腋花房の分化時期は11月はじめ頃であり、ハウス内の平均気温が摂氏25度以下になると促進されると考えられた。しかし、温度低下が緩やかなことから、花芽分化段階の個体間差が大きくなりやすいと考えられた。
4. 定植時の花芽分化段階および定植後の遮光の効果

第3表 定植時の花芽分化段階z
第4表 各処理区の頂花房~
第4次腋花房の出蕾・開花および収穫開始日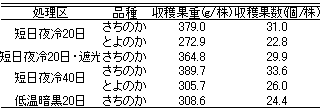
第5表 各処理区の4月末までの株当たりの
収穫果重および収穫果数
定植時の花芽分化程度(第3表)は、短日夜冷20日区の‘さちのか’で頂花房が肥厚後期~2分期、‘とよのか’で肥厚中期であった。40日区では、両品種ともに頂花房が雄ずい形成期~雌ずい形成期、第1次腋花房が肥厚中期であった。低温暗黒区では花芽分化は認められなかった。
頂花房の出蕾日(第4表)は、両品種とも短日夜冷40日処理区で最も早く、11月22日前後であった。短日夜冷20日処理区では、‘さちのか’で11月29日、‘とよのか’で12月6日であった。低温暗黒区では12月31日と大幅に遅れた。
4月末までの株当たりの収穫果重(第5表)が最も高かったは、‘さちのか’の短日夜冷20日区の379.0gで、ついで‘さちのか’の短日夜冷40日区の389.7gであった。最も少なかったのは、‘とよのか’の短日夜冷20日区の272.9gであった。株当たりの収穫果数は、‘さちのか’の短日夜冷40日区で33.6個と最も多く、次いで‘さちのか’の短日夜冷20日区の31.0個であった。最も少なかったのは、‘とよのか’の短日夜冷20日区の22.8個であった。
糖度は両品種とも8~12度の間で推移した(データ略)。酸度は、‘とよのか’の方が高く推移した。硬度は、全期間を通じて‘さちのか’の方が高く、3月以降も大きく低下することはなかった。
以上の結果から、‘さちのか’では花芽分化を誘導するために低温暗黒処理を行うより短日夜冷処理を行った方が収量が高くなる可能性が示唆された。また、‘さちのか’では3月以降の果実品質も良好で、収穫期間の延長も可能であると考えられた。
5. 花芽分化抑制処理の効果
定植時には、花芽分化抑制処理を行わない無処理区で第1次腋花房が分化、高温区で頂花房が分化しており、長日および高温長日区では花芽は未分化であった(データ略)。
頂花房の出蕾日(第6表)は、無処理区で11月9日と最も早かった。高温区では11月20日、長日および高温長日区では12月27日前後が頂花房の出蕾日となった。第1次腋果房以降は、各処理区とも1月ごとに出蕾が認められた。
4月までの株当たりの収穫果重(第7表)は、頂花房除去区で312.0gと最も重く、次いで高温区の301.2gであった。無処理区では268.7gと最低であった。収穫果数は、高温区で最も多かった。
以上の結果から、花芽分化を抑制するには長日および高温長日処理が有効であるが、沖縄では定植時に花芽が未分化であると花芽分化の遅れから収穫が大幅に遅れる可能性が考えられた。一方、花芽分化抑制処理を行わないと収穫は早まるが、早期の着果負担が大きく、全収量は抑えられた。頂花房を除去すると収穫果重、果数ともに大きく、着果負担が小さかったことから有望な方法であると考えられた。
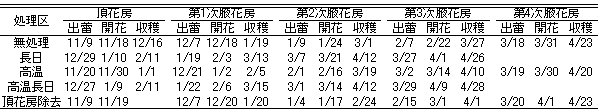
第6表 各処理区の頂花房~第4次腋花房の出蕾・開花および収穫開始日

第7表 各処理区の4月末までの株当たりの収穫果重および収穫果数
6. 最適な採苗時期と低温暗黒処理の効果

第8表 各処理区の定植時の
花芽分化程度zと分化率 
第9表 頂花房の出蕾に及ぼす採苗時期と
低温暗黒処理の影響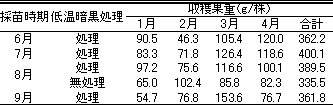
第10表 4月までの月別株当たりの収穫果重と
果実数に及ぼす採苗時期と低温暗黒処理の影響
定植時の花芽分化程度と分化率に及ぼす採苗時期の影響は明らかではなかったが、いずれの時期に採苗しても低温暗黒処理を行わない方が分化程度および分化率は高い値を示した(第8表)。
頂花房の出蕾日は、6月採苗・低温暗黒処理区と8月採苗・無処理区で早く、それぞれ11月29日および30日であった(第9表)。
7月および8月の採苗して低温暗黒処理を行った場合の頂花房の出蕾日は、それぞれ12月3日および5日であった。9月採苗・低温暗黒処理区では、12月9日に頂花房の出蕾が認められた。
4月までの株当たりの収穫果重(第10表)は、7月採苗・低温暗黒処理区で400.1gと最も高く、次いで8月採苗・無処理区の389.6gであった。最も低かったのは、8月採苗・低温暗黒処理区の335.5gであった。6月および9月採苗の低温暗黒処理区での1~2月の収穫果重は低かった。収穫果数についても同様の傾向が見られた。
以上の結果から、沖縄で6月に採苗すると育苗期間が長く、苗の老化を引き起こし、実用的ではないと考えられた。また、9月採苗では定植時の生育が劣り、1~2月の収量が低下した。そこで、沖縄での採苗時期は、収量の多かった7~8月が適すると考えられた。一方、低温暗黒処理による花芽分化誘導効果は明らかではなかった。
7. 沖縄におけるポット苗の頂花房分化時期
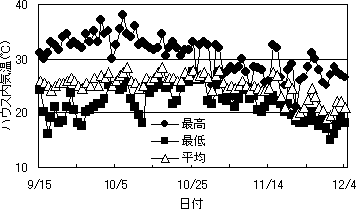
第4図 ハウス内気温の推移
沖縄県農試のサイドを開放した鉄骨ハウスでは、9月半ばに低温となる時期があったが、10月下旬になってもあまり気温は低下せず、日最低気温が摂氏20度を下回ったのは11月下旬であった(第4図)。
検鏡の結果、鉢上げ後約2週間後でも花芽分化している株があり、育苗中から出蕾する株もかなり見られた。一般的な肥培管理をした低N区で、本格的な花芽分化が始まったのは‘とよのか’で11月5日前後であった(第11表)。‘さちのか’では花芽分化株率がゆるやかに高くなり、50%を超えたのは11月10日以降であった。摂氏10度の低温暗黒処理を10日間行った場合の花芽分化の促進効果は明らかではなかったが、定植後の出蕾・開花は両品種ともに早まった。このことから、検鏡時には形態的の判定可能な状態には達していなかったが、生理的には分化が誘導されていたと推察された。
以上のように、冬季に低温遭遇が不十分な沖縄では、生産株から採苗した場合には育苗中から開花する
株も多く、頂花房の分化段階が大きくばらつくことが明らかとなった。また、頂花房の分化時期は、気象経過、すなわち年次により大きく変化すると考えられた。

第11表 各処理区の花芽分化程度zと分化株率の推移
8. 沖縄に適した短期栽培における作型の検討
2000年の秋季は非常に高温で経過したため、ポット苗の自然分化時期(第12表)は‘さちのか’では11月10日頃と判断された。沖縄における生産株から得られた苗では、育苗期から出蕾・開花する株が多く、10月31日頃では2~3割程度であった。低温暗黒処理による花芽分化の促進効果は明らかではなかった。今回最も遅く定植したのは10月31日であったが、この時期でも大半の株で花芽分化は認められなかった。低温暗黒処理を行った区では、検鏡による花芽分化段階の差は明らかではなかったが、定植後の出蕾・開花は明らかに早まり、年内から収穫が始まった。
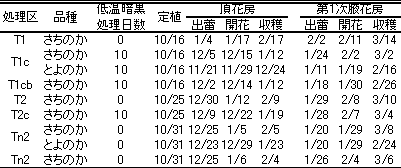
第12表 各処理区の頂花房と第1次腋花房の出蕾・開花および収穫開始日
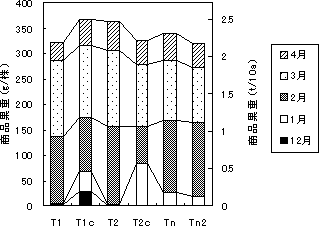
第5図 各処理区の‘さちのか’の月別収量
株当たりの収量(第5図)はT1c区で最も多く、3月末までで320g(2.2t/10a)、4月末までで370g(2.5t/10a)であった。平均1果重は収穫初期には大きかったが、3~4月になると10g前後であった。収量は、‘とよのか’より‘さちのか’で高かった。
12月の果実の糖度は8~9度であり、酸度は1%とやや高かったが、その後0.8%程度まで低下した。果実品質は時期的な変動が大きく、収穫直前の気象条件にかなり左右されていると思われた。
第1次腋花房の芽数を2芽にしたTn2区でも、特に収量が増加することはなかった。花房の上にネットで遮光したTcb区でも、果実品質の向上効果は認められなかった。以上の結果から、12月20日頃に充分な収量を得るためには、10月10日頃の定植が目安となり、頂花房の花芽分化処理が不可欠であると考えられた。
9. 沖縄でのイチゴ栽培に適した花芽分化処理法の検討
A. 低温暗黒処理による花芽分化促進効果

第6図 11月1日での各処理区の花芽分化程度
低温暗黒処理を終了した11月1日では、低N・15日および10日区ですべての株が花芽分化していた(第6図)。高N・10日区では、低N・10日区に比較してが分化がやや遅れるとともにその分布は不均一であり、未分化の株も認められた。低N・20日および高N・20日区では、それぞれ70および80%の株が未分化であった。
以上のように、育苗中のN施用量は少ない方が花芽分化は促進され、その斉一性は高いと考えられた。一方、低温暗黒処理期間は長い方が花芽分化は逆に抑制される傾向があったことから、沖縄では低温暗黒処理による花芽分化促進効果は低く、有効な花芽分化処理法ではないと考えられた。
B. 沖縄でのイチゴ栽培に適した花芽分化処理法
無処理区では10月16日になって花芽分化した株が認められたが、半数以上の株で未分化であった。生産株から採苗するより休眠打破株から採苗した方が、わずかに花芽分化は進んでいた(第7図)。短日夜冷36日区では、処理開始16日後の9月26日に10%の株で花芽分化が認められた。10月5日になると、すべての株で花芽分化しており、分化段階は肥厚中期~がく片形成期になった。10月16日では、すべての株の頂花房が雄ずい形成期にあり、第1次腋花房も肥厚初期~肥厚後期にあった。短日夜冷20日区では、処理開始15日後の10月11日にすべての株で花芽分化が確認され、分化段階は肥厚初期~がく片形成期にあった。10月16日の花芽分化段階はがく片形成期~雄ずい形成期であった。
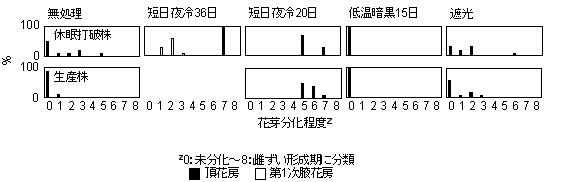
第7図 定植時(10月16日)での各処理区の花芽分化程度
定植2週間後の調査では、採苗した親株に関係なく、低温暗黒15日および遮光区で葉数が少なく、クラウン径が小さくなる傾向が見られた(データ略)。これらの処理区では受光量が少なくなり、光合成量が低下することによって、定植時に株が小型するものと考えられた。
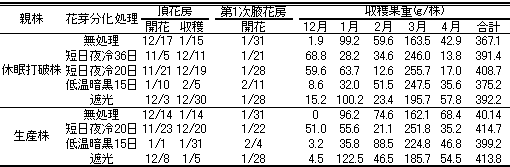
第13表 頂花房と第1次腋花房の開花・収穫開始日と収穫果重に及ぼす花芽分化処理法の影響
休眠打破株から採苗した場合、無処理区での頂花房は12月17日に開花し、1月15日に収穫が開始した(第13表)。遮光区では、これよりそれぞれ2週間程度早まった。短日夜冷36日区では、開花・収穫が35~40日早まり、20日区でも25日程度早まった。一方、低温暗黒15日区での開花・収穫は、無処理区より20日以上遅れた。第1次腋花房は、無処理区では1月31日に開花した。遮光区では1月28日に開花し、短日夜冷36日および20日区の開花日はそれぞれ1月21日および28日であり、短日夜冷処理による第1次腋花房の開花促進効果は低かった。低温暗黒15日区での第1次腋花房の開花日は2月11日であり、無処理区より10日程度遅れた。生産株から採苗した場合にも、各区の頂花房の開花・収穫開始および第1次腋花房の開花は、休眠打破株から採苗した場合とほぼ同様であった。4月末までの収量(第13表)は、休眠打破株および生産株から採苗した場合ともに、低温暗黒15日区で低くなった。他の試験区間差は明らかではなった。休眠打破株から採苗した場合、無処理、低温暗15日および遮光区でも12月に収穫が始まったが、収量はごくわずかであった。短日夜冷36日および゙20日区では、12月に十分な収量が得られ、36日区でやや収量は高かった。しかし、36日区では1月の収量が少なくなった。低温暗黒15日区でも1月の収量は少なく、そのため全体的な収量も低くなった。短日夜冷20日区では2月の収量が低くなった。生産株から採苗した場合、短日夜冷20日区でのみ年内収量が得られた。1月の収量は低温暗黒15日区で低く、2月の収量は短日夜冷20日区で低かった。
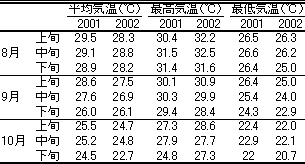
第14表 沖縄地方(金武町)での
2001および2002年の8~10月の気温の推移
2002年の試験では、定植時に無処理および遮光区でもすべての株で花芽分化が認められた(データ略)。低温暗黒15日区ではほとんどの株が未分化であった。短日夜冷20日区の花芽分化程度は無処理および遮光区と大差なかったが、分化程度のばらつきは小さかった。2002年の8月下旬以降の最低気温(金武町)は、2001年より摂氏0.5~1.5度低く推移した(第14表)。このため、無処理および遮光区でも花芽分化が早まり、短日夜冷20日区の効果は明らかではなかったと考えられた。
沖縄では低温暗黒処理は有効な花芽分化処理法ではなく、10月中旬定植では短日夜冷処理が効果的であった。また、花芽分化に及ぼす親株の前歴の影響は小さいものと考えられた。短日夜冷処理を36日間行うと定植時の花芽分化程度は進むが、20日間処理と年内収量の差は少なく、1月の収量は20日間処より少なくなった。従って、十分な年内収量を確保するためには、20日間程度の短日夜冷処理を行った苗を10月中旬に定植するのが有効であると考えられた。
以上の結果から、沖縄での低温暗黒処理は花芽分化を遅らせ、10月中旬定植での安定的かつ効果的な花芽分化処理法は短日夜冷処理であり、20日間の処理で頂花房を分化させることができた。また、親株の前歴による花芽分化あるいは収量への影響は小さいと考えられた。
10. セル育苗の可能性と定植後の水分管理法の検討

第15表 株当たりの月別収穫果重に及ぼす灌水管理と育苗方法の影響
いずれの育苗方法でも定植時に頂花房~第1次腋花房が分化していたが、セルトレイで育苗した場合には花芽分化が遅れ、その分布のばらつきは大きかった(データ略)。
灌水直後のpF値は灌水処理に関係なく1.2~1.3程度であったが、通常灌水区では2.0前後、少灌水区では2.3~2.4前後まで上昇した(データ略)。
セルトレイおよび7.5cmポットで育苗した場合、定植時には葉数が少なく、クラウン径も小さくなり、株が小型化する傾向が見られた(データ略)。その傾向は定植1ヶ月後も見られたが、2ヶ月後になると差は見られなかった。一方、灌水処理による明確な生育差は見られなかった。
4月末までの株当たりの収穫果重(第15表)に及ぼす灌水処理による影響は明らかではなかった。セルトレイで育苗した場合には1月の収量が低下した。これは定植時の花芽分化と生育の遅れが関係すると考えられた。一方、10.5cmポットと7.5cmポットで育苗した場合の月別および全体的な収量の差は明らかではなかった。
以上の結果から、7.5cmポットおよびセルトレイで育苗した場合、従来の10.5cmポットでの育苗に比べ、定植時の苗が小さくなり、その影響は定植1ヶ月後も続いた。セルトレイで育苗すると花芽分化はやや遅れ、初期収量も減少した。全体的な収量から見ると、7.5cm程度のポットでの育苗は可能であると考えられた。一方、灌水処理による生育・収量への影響は明らかではなかった。本年は、沖縄では1~3月の雨量が少なかったことが、このことに関係している可能性も考えられた。
11. 定植後の水分・施肥管理法の検討
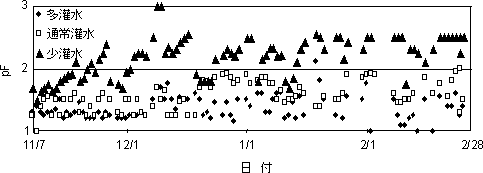
第8図 2002年11月17日~2003年2月28日までのpF値の推移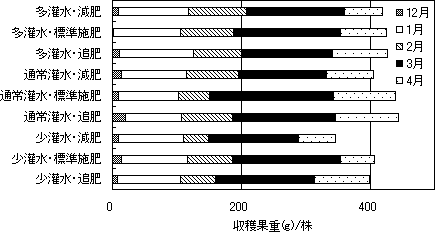
第9図 株当たりの月別収穫果重に
及ぼす灌水・施肥管理の影響
pF値は概ね、多灌水区では1.2前後、通常灌水区では1.5~2.0,少灌水区では2.5前後で推移した(第8図)。
2月末の葉数、新生第3葉の葉柄長、葉身長、葉幅および葉色、ならびにクラウン径のいずれについても、灌水・施肥管理の影響は見られなかった(データ略)。また、葉柄の搾汁液の硝酸イオン濃度は減肥区で低く、追肥区で高くなったが、カリウムイオン濃度については施肥管理の影響は明らかではなかった。灌水管理の影響は、硝酸およびカリウムイオンともに明確ではなかった。
灌水量、施肥量が少なくなる方が、4月末までの株当たりの収穫果重は少なくなる傾向が見られた(第9図)。ただし、多灌水での減肥による収量の低下は小さかった。
以上のことから、灌水・施肥量が少なくなると生育への影響は小さくとも、収量が低下し、高収量を上げるためには適度な灌水と追肥が必要であると考えられた。
エ. 今後の課題
当初は、1~2月に収穫する短期作型の開発を目標としてきたが、‘さちのか’を用い、短日夜冷処理により花芽分化を誘導した苗を10月中旬に定植することにより、12~4月の収穫が可能となった。収量についても、2~3t/10aは達成できると考えられる。しかし、沖縄では秋~冬季の気象変動が大きく、また、土壌条件についても本土とは異なる。採苗時期、定植時期、花芽分化処理や定植後の灌水・施肥管理については、一定の成果が得られたが、現地にイチゴ栽培を導入し、安定的な生産を行うために、今後も細部にわたる個別の試験研究が必要であると考えられる。
オ. 要約
沖縄では冬季もサイドを開放した簡易ハウスでイチゴの栽培が可能であり、暖房、電照、ジベレリン処理は不要であった。品種については、収量・品質面から、‘とよのか’より‘さちのか’が適すると考えられた。‘さちのか’では果実が硬いことからも沖縄での栽培に適した。採苗時期については、6月では苗の老化から、9月では定植では定植時の生育の遅れから収量が低下し、収量の多かった7~8月の採苗が適当であると考えられた。自然状態で花芽分化した苗を定植する場合、11月1日頃が定植適期であると考えられた。ハウス内気温が摂氏25度以下になると花芽分化は促進され、分化時期は頂花房が10月20日頃、第1次腋花房が11月はじめ頃と考えられた。しかし、温度低下が緩やかなであるため花芽分化段階の個体間差が大きく、また年次間の分化時期にも差が生じると考えられた。花芽分化した苗を10月中旬定植すると、年内からの収穫が可能であった。その場合の花芽分化処理法は、低温暗黒処理法では効果がなく、短日夜冷処理が有効であった。20日間の短日夜冷処理で頂花房が、40日間の処理で第1次腋花房が分化するが、収穫開始時期については差が小さく、40日間の処理では1~2月の収量が低下することから、短日夜冷処理の期間は20日間程度が適当であると考えられた。また、セルトレイを用いて育苗すると、花芽分化が不安定であり、定植時の生育が遅れることから、初期収量が低下した。このため、育苗には10.5cm程度のポットを用いるのが適当であると考えられた。定植後の灌水および施肥量が減少すると、収量低下につながり、特に灌水量の影響は大きいと考えられた。
カ. 文献
- 高市益行・田中和夫・中島規子・野口裕司・宮城信一・久場峯子・山崎篤・荒木陽一、1999. 亜熱帯(沖縄)における促成イチゴの短期栽培に適した苗の定植時期・花芽分化段階.園芸学会九州支部研究集録.7:49.
- 高市益行・田中和夫・中島規子・野口裕司・宮城信一・久場峯子・山崎篤・荒木陽一、1999. 亜熱帯(沖縄)における促成イチゴ品種‘とよのか’および‘さちのか’の開花・生育特性.園学雑.68(別2):254
- 中島規子・宮城信一・高市益行・久場峯子・荒木陽一、2000. 沖縄におけるイチゴ花芽の自然分化期と温度の関係.園芸学会九州支部会研究集録.8:71.
- 中島規子・宮城信一・高市益行・久場峯子・荒木陽一、2000. 沖縄における採苗時期の違いと株冷処理の有無がイチゴの花芽分化・生育および収量に及ぼす影響.園学雑.69(別2):341.
- 高市益行・登野盛博一・中島規子・荒木陽一・小橋川共志・久場峯子、2001. 亜熱帯の沖縄におけるイチゴ品種‘とよのか’、‘さちのか’の花芽分化特性および収量.園学雑.70(別2):296.
- 大和陽一・中島規子・高市益行・登野盛博一・小橋川共志・久場峯子・喜納兼二・荒木陽一・山口博隆、2002. 沖縄地方でのイチゴ栽培に適した花芽分化処理法.園学雑71. (別2):371.
- 九州沖縄農業研究センター野菜花き研究部耐暑性野菜生産研究室、2003. イチゴ(2)-沖縄に適した栽培技術(花芽分化処理を中心として)-.p.8-9. 地域総合研究「亜熱帯」総括事務局編.島を興す-農の新技術-新規野菜・花き栽培技術マニュアル.