背景・経緯
多くの越冬性植物は晩秋の低温にさらされると、冬の凍結温度にも耐えて越冬できる能力を獲得します。この現象を低温馴化と呼びますが、どの様な仕組みで耐凍性が獲得されるのかについては、よく分かっていません。低温馴化のメカニズムを解明することにより、植物の耐凍性が何によって決められているのか耐凍性を強化するにはどの様にすべきかなどが明らかになると考えられています。本研究では、低温馴化の過程で植物が蓄積するタンパク質の1つに注目しました。このタンパク質は、細菌で低温ショックタンパク質として知られているタンパク質と同じ構造と機能(RNAシャペロンと呼ばれるRNA構造変換活性)をもっていることが明らかになりました。更にRNAシャペロンタンパク質が植物の耐凍性を制御していることを世界で初めて明らかになりました。
内容・意義
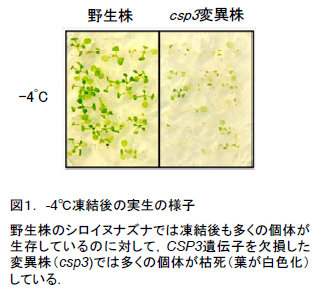
- シロイヌナズナのもつ低温ショックタンパク質の1つであるCSP3タンパク質を作り出す遺伝子を欠失させると、耐凍性が著しく低下していることが明らかになりました(図1)。また、この植物では、ストレス耐性に関与するいくつかの遺伝子の発現が低下していました。
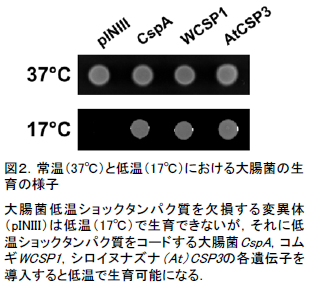
- 低温ショックタンパク質を欠損した大腸菌は低温環境下では生育できなくなります。この大腸菌にCSP3遺伝子を導入し、CSP3タンパク質を作らせると、低温でも生育が可能になりました。つまり、シロイヌナズナのCSP3タンパク質は大腸菌の低温ショックタンパク質と同じくRNAシャペロン機能を持っていることがわかりました(図2)。
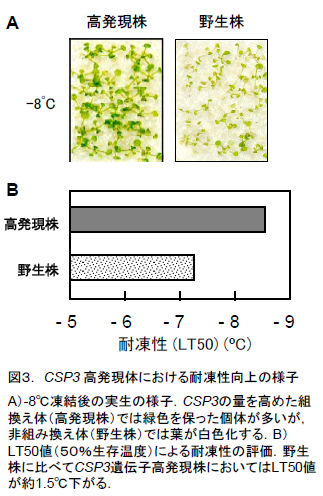
- CSP3遺伝子を高発現させ、その働きを常に高めるように組換えたシロイヌナズナでは耐凍性が向上しており、耐性限界温度(LT50)が-7.2°Cである野生型(非組み換え体)より約1.5°C下げることに成功しました(図3)。このとき、CSP3遺伝子の高発現組換え体は、野生型と同じように生育し、生育に対する負の影響は見られませんでした。
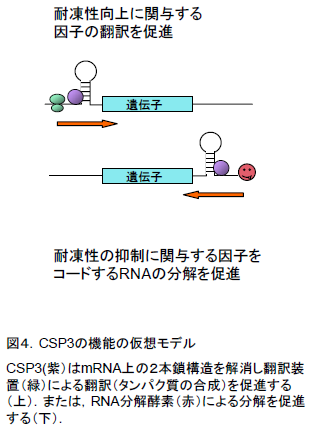
- CSP3タンパク質はRNAシャペロン活性により、耐凍性に関わる遺伝子をコードするmRNAの翻訳や分解を調節していると予想されます(図4)。
本研究では、これまでに知られていなかった新しい機構で植物の耐凍性がコントロールされていることが明らかになりました。これは、RNAシャペロン活性が植物の環境適応の制御に関与していることを示した点で画期的な知見です。
今後の予定・期待
CSP3と同様の遺伝子は、ほぼ全ての植物に見出されます。これらの遺伝子を高発現させることで、コムギ、牧草などの越冬性の向上や野菜、果樹への霜害抵抗性付与などが可能になると思われます。
参考データ
用語の解説
【耐凍性】
0°C以下の凍結温度に対する耐性。細胞内の水分が凍結すると枯死に至るので、越冬性の植物には、様々なタンパク質や糖類を細胞内に蓄積して、マイナス温度でも凍結しないような仕組みが存在する。
【低温ショックタンパク質】
大腸菌などのバクテリアにおいて、低温で誘導されるタンパク質で、低温ショックドメインとよばれる構造を共通に持つ。大腸菌ではRNAシャペロン活性を持つ。
【RNAシャペロン】
通常は一本鎖(直鎖)であるRNA中に生じた部分的な2本鎖(ヘアピン)構造に結合し、1本鎖構造に戻す活性を持つタンパク質。
【耐性限界温度(LT50)】
50%の個体が死滅する温度。耐凍性を示す指標として用いられる。