研究テーマについて
キク矮化病はキク矮化ウイロイド(Chrysanthemum Stunt Viroid, CSVd)がキクに感染することによって発生します。キクに感染すると草丈がわい化します。このようなキクは商品価値がほとんどありませんので生産者にとっては非常に頭の痛い問題です。CSVdがキクに1度感染すると自然に消滅することはありませんので廃棄するしかありません。また、感染株は他の健全株への汚染源になる可能性があります。
キク矮化病は1977年に国内での発生の報告があり、それ以降各地で発生報告がされており、ほぼ全国のキク産地で発生していたことが確認されています。
以上のように、キク矮化病の発生は重要な問題です。そこで本研究所ではキク矮化病の発生生態の解明と診断に取り組んでいます。
 わい化病発病個体と健全個体 写真の中央にあるキクが発病個体です。 周囲にある健全個体と比較して半分程度になっています。 |
 CSVdに感染したミスルトーの葉 キク品種‘ミスルトー’はCSVdに感染すると葉に 退緑斑点が生じることが知られています。 生物検定用の品種として用いられています。 |
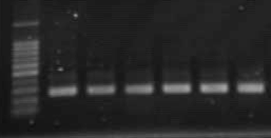
RT-PCRで得られた産物の泳動写真
(CSVdの検定結果)
写っているバンドが増幅された産物です。
CSVdの検定方法は生物検定・PAGE・ハイブリダイゼーション・RT-PCRなどがあります。本研究所では主にRT-PCRによってCSVdの検定を行っています。検定方法はサンプル(葉・茎・根など)に針など(先の尖ったものであれば何でもよい)を挿し、それを逆転写溶液に浸して逆転写反応を行ないます。そのあとは一般的なRT-PCRの操作と同じです(詳細はJournal of Virological Methods,2006, 131( 1) 28-33)。いずれの方法を用いる場合であっても、検定自体が正確に行われていることを確認するために、検定の際には陽性となるサンプルが必要です 。
一部の品種では冬季の低温遭遇によってCSVdの濃度が低下することが知られています。そのため、RT-PCRによる検定においても検出できなくなることがあります。実際に、低温状態においては植物体内におけるCSVdの感染分布に偏りが生じることが確認されています。

CChMVdに感染したキクの葉
近年、国内での発生が確認されたウイロイド病です。
症状は葉の緑色が薄くなり、場合によっては退緑斑を生じます。
キクには様々な品種があることが知られていますが、その中にはわずかに抵抗性品種があることが最近知られるようになりました。その抵抗性は後代へ遺伝することが判明していることが、今後のキク矮化病防除に役に立つ形質であると考えられています。
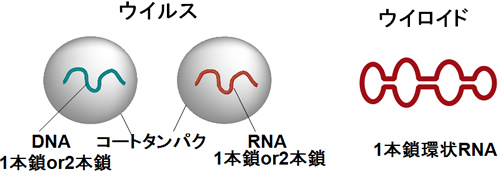
ウイロイド(viroid)
ウイロイド(viroid)とは1本鎖環状RNAから成る最小の植物病原体(250~400ヌクレオチ ド)であり、1971年にDienerらによってジャガイモから発見されました。国内ではキク以外に、カンキツ、ホップ、リンゴなどで発生の報告があります。ウイロイドRNAはタンパク質をコードする遺伝子が刻み込まれていないことが明らかになっており、ウイルスのようなコートタンパクを持たず、また、ウイロイドRNAの複製は宿主細胞の酵素を利用していると考えられています。ウイロイドは塩基配列相同性に基づいた分類によると、 ポスピウイロイド科(Pospivioridae)とアブサンウイロイド科(Avsaunviroidae)の2つに分けられています。ポスピウイロイド科は中央保存領域と呼ばれる保存性の高い領域があ り、アブサンウイロイド科は中央保存領域を持たないが、ハンマーヘッド型リボザイム活性を有し、自己切断するという特徴を持つとされています。また、前者は核内で増殖し、後者は葉緑体内で増殖するとされています。
キクに感染するウイロイドはキク矮化ウイロイド(Chrysanthemum stunt viroid, CSVd)と(Chrysanthemum chlorotic mottle viroid, CChMVd)が知られており、前者はポスピウイロイド科、後者はアブサンウイロイド科に属します。
※日本植物病理学会・植物ウイルス分類委員会作成の「日本に発生する植物ウイルス・ウイロイド (2014)」によってCSVdは「キク矮化ウイロイド」、CChMVdは「キク退緑斑紋ウイロイド」と日本語名が決定しました。
ウィルスとウイロイド
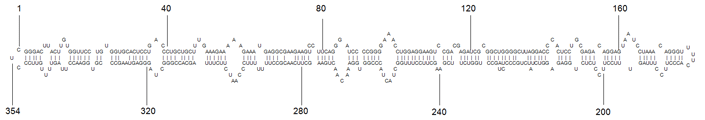
代表的なキク矮化ウイロイド(CSVd)の二次構造図
参考文献
- 佐野輝男 ウイルス 60, 177-186(2010)
- 松下陽介 農業技術大系.花卉編6.キク(クリサンセマム).ウイルス、ウイロイド 農山漁村文化協会 (2008)
主な研究成果
- 松下陽介 2016. ウイロイド~起源・伝播・進化~ 化学と生物 54(3), 170-175.
- Y. Matsushita*, Y. Shima. 2015. Effect of low temperature on the distribution of Chrysanthemum stunt viroid in Chrysanthemum morifolium Phytoparasitica 43, 609-614.
- 浅野 俊介、松下 陽介、平山 喜彦、仲 照史 2013. ダリアに感染するウイルス・ウイロイドの検出技術の開発および国内における発生状況 植物防疫 69(12), 12-16.
- S. Asano, Y. Matsushita*, Y. Hirayama, T. Naka. 2015. Simultaneous detection of Tomato spotted wilt virus, Dahlia mosaic virus, and Chrysanthemum stunt viroid by multiplex RT-PCR in dahlias and their distribution in Japanese dahlias. Letters in Applied Microbiology 61, 113-120.
- Y. Matsushita. 2013. Chrysanthemum stunt viroid. Japan Agricultural Research Quarterly (JARQ) 47, 237-247.
- 松下陽介 2013. キク矮化ウイロイド抵抗性の遺伝と選抜 植物防疫 67(6), 33-35.
- Y. Matsushita*, K. Aoki, K. Sumitomo. 2012. Selection and inheritance of resistance to Chrysanthemum stunt viroid. Crop Protection 35, 1-4.
関連する成果情報 → 「キク矮化ウイロイド抵抗性品種の選抜と抵抗性の後代への遺伝」 - Y. Matsushita, P. K. R. Kumar*. 2009. In vitro transcribed Chrysanthemum stunt viroid (CSVd) RNA is infectious to Chrysanthemum and other plants. Phytopathology 99, 58-66.
- Y. Matsushita*, T. Tsukiboshi, Y. Ito, Y. Chikuo. 2007. Nucleotide Sequences and Distribution of Chrysanthemum Stunt Viroid in Japan. Journal of the Japanese Society for Horticultural Science 76, 333-337.