ゲノム編集技術による穂発芽耐性の改良されたコムギ育種素材
要約
2倍体であるオオムギの遺伝子情報と配列特異的に変異を導入できるゲノム編集技術を用いて、重複するコムギの標的遺伝子を一度に改変することで作出した育種素材である。穂発芽耐性を改良したコムギ品種の開発が期待できる。
- キーワード:穂発芽、ゲノム編集、コムギ、高次倍数性、CRISPR/Cas9
- 担当:次世代作物開発研究センター・基盤研究領域・育種法開発ユニット
- 代表連絡先:電話 029-838-7930
- 分類:研究成果情報
背景・ねらい
コムギの収穫時期は梅雨の季節と重なり、穂発芽による被害がしばしば発生する。コムギの穂発芽耐性育種の歴史は長いが、その耐性は十分でなく、さらなる改良が望まれている。穂発芽耐性には種子休眠性が深く関与しており、近縁で2倍体作物のオオムギにおいて、遺伝子機能が欠失することで種子休眠が長くなるQsd1遺伝子(quantitative trait locus on seed dormancy 1)が同定されている。しかし、コムギは異質6倍体で同祖の遺伝子を重複して3組持っていることもあり、Qsd1同祖遺伝子の座乗位置に種子休眠性との関係を示す遺伝解析に関する報告はこれまでない。そこで、本研究では、配列特異的に変異を導入することのできるゲノム編集技術を活用して、コムギの3つのQsd1同祖遺伝子(TaQsd1-A, B, D)の機能を一度に欠失させることで、穂発芽耐性の改良された育種素材を作出できるのか明らかにすることを目的とする。
成果の内容・特徴
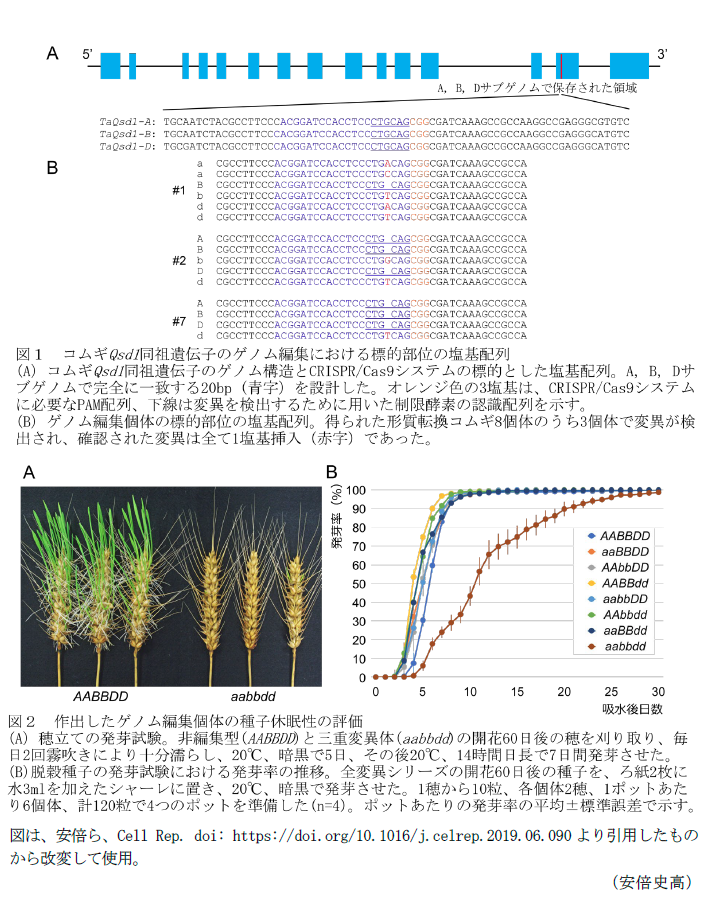
- CRISPR/Cas9システムの標的配列(図1A)を用いて、アグロバクテリウムを介したコムギ形質転換法により得たゲノム編集個体の中には、TaQsd1-A, B, D全てに同時に変異の導入されている植物体が含まれる(図1B、#1:遺伝子型aaBbdd)。
- TaQsd1-A, B, D全てに変異を導入した植物体(aaBbdd)と野生型(AABBDD)との交配を行うことで、ゲノム編集のために用いた外来遺伝子を遺伝分離で除去し、かつ変異をホモ固定した全8組合せの変異シリーズをF2世代で一度に得ることができるので、倍数性のコムギにおける変異体の作出にかかる期間を従来の手法に比べて大幅に短縮できる。
- 霧吹きにより雨濡れ状態を再現した穂立ての発芽試験において、非編集型(AABBDD)に比べて三重変異体(aabbdd)は明らかに発芽が抑えられる(図2A)。また、全ての変異シリーズについて脱穀して発芽試験を行った場合、7日目の発芽率の平均が三重変異体以外では80%以上であるのに対して、三重変異体は約20%であり、三重変異体のみ有意に種子休眠が長い(図2B)。
- 三重変異のゲノム編集個体、野生型、外来遺伝子を保持しているT1個体について、次世代シーケンサーを用いてコムギゲノム(16Gb)の30倍量以上にあたる塩基配列情報を取得して比較解析すると、ゲノム編集個体と野生型に違いは見られない。
成果の活用面・留意点
- コムギの穂発芽耐性を改良させる新規の遺伝資源としての活用が期待される。
- コムギにおけるQsd1同祖遺伝子変異の効果は、形質転換が可能なコムギ品種「Fielder」の遺伝背景で得られる結果であり、他品種の遺伝背景での効果は個別に確認する必要がある。また、閉鎖系の人工気象室で育成する場合の結果であり、様々な環境要因が影響する圃場においては、発芽試験の温度などの複数の条件で複数年試験するなど総合的な評価も必要である。
- コムギではQsd1同祖遺伝子の三重変異で初めて形質が現れることが実証され、このことは、より単純なゲノム構造を持つオオムギの遺伝子情報を活用したゲノム編集技術が、高次倍数性作物のコムギにおいて特に有用となることを示している。他の形質、遺伝子での応用が期待される。
具体的データ
その他
- 予算区分:交付金、競争的資金(科研費)、その他外部資金(SIP)
- 研究期間:2014~2019年度
- 研究担当者:
安倍史高、ハク エムダドウル、久野裕(岡山大)、田中剛、神谷容子(横浜市大)、三上雅史、川浦香奈子(横浜市大)、遠藤真咲、大西一光(帯広畜産大)、林武司、佐藤和広(岡山大) - 発表論文等:Abe F. et al. (2019) Cell Rep. 28:1362-1369