米でんぷんの構造を制御する遺伝子座とアルカリ崩壊性遺伝子座の連鎖
※アーカイブの成果情報は、発表されてから年数が経っており、情報が古くなっております。
同一分野の研究については、なるべく新しい情報を検索ください。
要約
主にジャポニカ品種とインディカ品種間にみられる米でんぷんのアミロペクチン構造の違いは、一つの主働遺伝子によって制御されている。その染色体上の位置は、米でんぷんの糊化特性に関わるとされるアルカリ崩壊性遺伝子座に近い。
- 担当:東北農業試験場・水田利用部・栽培生理研究室
- 連絡先:0187-66-1221 ume@omg.affrc.go.jp
- 部会名:水稲
- 専門:栽培
- 対象:稲類
- 分類:研究
背景・ねらい
最近、イネ胚乳でんぷんのアミロペクチンには、構造が明確に異なる2つのタイプの存在が明らかとなった(平10成果情報、Umemoto et al.,1999)。この違いはでんぷんの特性、ひいては米の食味、加工適性等に影響を及ぼす可能性がある。これを証明するには、アミロペクチン構造のみが異なる準同質遺伝子系統を選抜、育成し、食味・加工適性試験等を行う必要がる。しかし、選抜にはアミロペクチン構造を判定する簡便な手法が求められる。
これまでの知見より、アミロペクチン構造の変異は、イネ胚乳のアルカリ崩壊性の原因である可能性がある。そのため本研究により両形質を制御する遺伝子座間の強い連鎖が証明されれば、検定の容易なアルカリ崩壊性の調査により、目的とするアミロペクチン構造を持つ系統の選抜が可能となる。
成果の内容・特徴
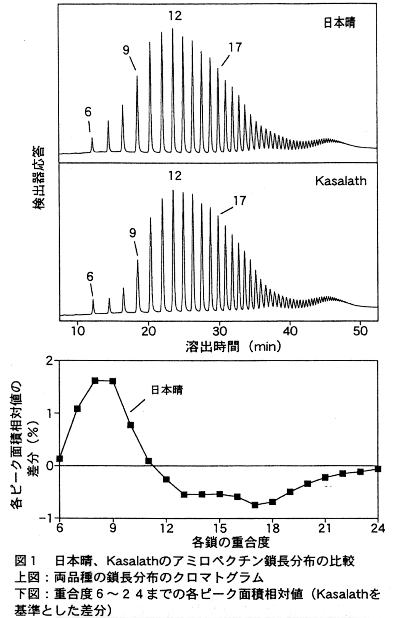
- 日本晴とKasalathのアミロペクチン鎖長分布には明確な差がみられ、日本晴は Kasalathより短鎖(重合度6~11)が多く、中鎖(12~23)が少ない(図1)。
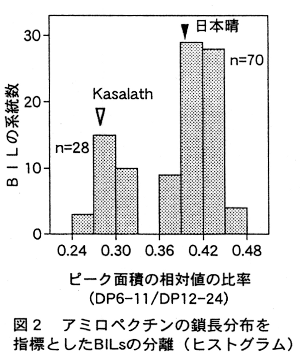
- 日本晴とKasalathの戻し交雑後代自殖系統群(BILs)のアミロペクチン鎖長を分析し、短鎖/中鎖の比を指標として分類したところ、日本晴型とKasalath型に分離し、その分離比から1つの主働遺伝子が関与している(図2)。
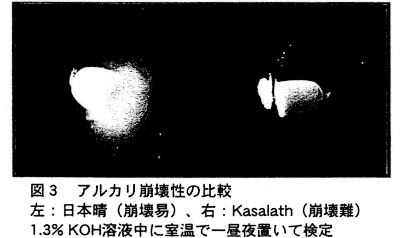
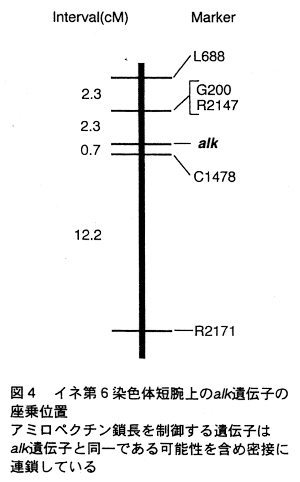
- アルカリ崩壊性(alk)についても分析を行い(図3) 、鎖長を制御する遺伝子とともに遺伝子座を決定した(図4)。両遺伝子ともに第6染色体短腕上の、極めて接近した位置に座乗することが明らかとなり、両遺伝子が同一である可能性も含めて、両者は強く連鎖している。
成果の活用面・留意点
- アルカリ崩壊性を利用した簡易検定法により、アミロペクチン構造の異なる系統の選抜が可能である。
- アルカリ崩壊性は、同一品種でも登熟期の温度によって変化することが知られている。また、ジャポニカ品種間のアルカリ崩壊性の違いは、alk遺伝子以外の要因も考えられる。
具体的データ
その他
- 研究課題名:イネ胚乳のアミロペクチン構造を制御する遺伝子座とアルカリ崩壊性 遺伝子座(alk)の連鎖解析
- 予算区分:経常(場特定)
- 研究期間:平成11年度