開発の社会的背景
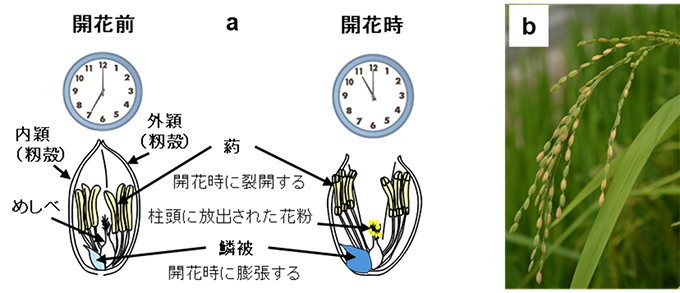
2023年、国連のアントニオ・グテーレス事務総長は「地球沸騰化時代」という言葉を用い、気候変動への危機感を露わにしました。実際に近年の異常高温は、様々な作物生産に影響を及ぼしつつあります。イネが高温に対する感受性が最も高いのは開花期で、35° C以上の高温下で開花すると、葯(やく)4) が柱頭(ちゅうとう)5) に花粉を放出できないため、受粉の失敗によりコメが実らない不稔(コメが実らない空籾)が増加します(図2 参照)。現在のところ、高温不稔による収量低下は大きな問題にはなっていませんが、イネは日本のみならず世界の約半分の人口にとっての主食であることや、異常高温が常態化してきている状況から、開花時の高温不稔を軽減できるイネは世界的にも重要な開発目標となっています。
図2 イネの正常な開花の模式図(a)と不稔が発生した穂(b)
研究の経緯
イネの通常の開花時刻である午前10時から12時は、日中の最高気温に向けて気温上昇している時間帯にあたります。この暑い時間帯での開花を避け、気温の低い早朝に開花する「早朝開花イネ」を開発できれば、高温不稔を軽減できることはすでに知られていました。本研究グループはこれまでの研究から、イネの第3染色体上に早朝開花性に関与する遺伝子が存在する可能性を示唆する結果を得ていましたが、どの遺伝子がどのような仕組みで開花時刻を早めているのか、詳しいメカニズムは分かっていませんでした。そこで本研究では、開花時刻を決定する遺伝子の特定と、早朝開花を引き起こす塩基配列の変異の同定を目指しました。
研究の内容・意義
① 早朝開花性の原因遺伝子の特定に成功
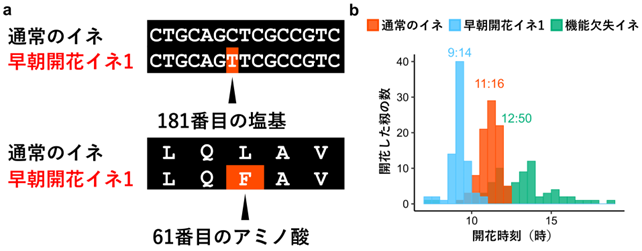
本研究グループが独自に育成した早朝に開花するイネと通常のイネの塩基配列を比較し、第3染色体上にある一つの遺伝子(Os03g0145400)が開花時刻に影響を及ぼしていることを突き止めました。本研究グループはこの遺伝子を「Early Morning Flowering 3 (EMF3 )」と命名しました。早朝開花イネ1ではEMF3 遺伝子の塩基配列のうち、181番目の塩基がシトシン(C)からチミン(T)に置換され、塩基の置換に伴いEMF3タンパク質の61番目のアミノ酸がロイシン(L)からフェニルアラニン(F)に変化していました(図3a )。EMF3 は2172の塩基配列から構成されますが、そのうちのたった1塩基の置換に起因する1アミノ酸の違いがイネの早朝開花性に関与すると考えられました。さらに、通常のイネでは午前10時から12時に集中して開花しますが、人為的にEMF3タンパク質の機能を欠失させたイネでは早朝から夕方までの幅広い時間帯に分散して開花するようになりました。この結果はEMF3 遺伝子がイネの開花時刻に大きな影響を及ぼすことを示します(図3b )。
図3 早朝開花イネ1の原因となる塩基やアミノ酸の変化 EMF3 遺伝子は2172の塩基から構成されますが、図3aでは181番目の塩基周辺の配列のみを示しています。通常のイネは181番目の塩基がシトシン(C)ですが、本研究グループが独自に育成した早朝開花イネ1ではチミン(T)に置換されていました。その結果、EMF3タンパク質の61番目のアミノ酸がロイシン(L)からフェニルアラニン(F)に変化します。EMF3 遺伝子に変異のあるイネの開花時刻。図中の時刻はそれぞれのイネの開花が最も盛んな時刻(開花時刻のピーク)を表しています(以下の図4 、5 も同様です)。通常のイネと早朝開花イネ1ではそれぞれ11:16と9:14に開花時刻のピークが見られますが、人為的にEMF3タンパク質の機能を欠失させたイネ(機能欠失イネ)では開花時刻のピークが緩やかで、早朝から夕方までの幅広い時間帯に開花がみられました。沖縄県石垣市での結果。
② EMF3 遺伝子に特定の変異を有する早朝開花イネでは開花時の高温不稔が軽減される
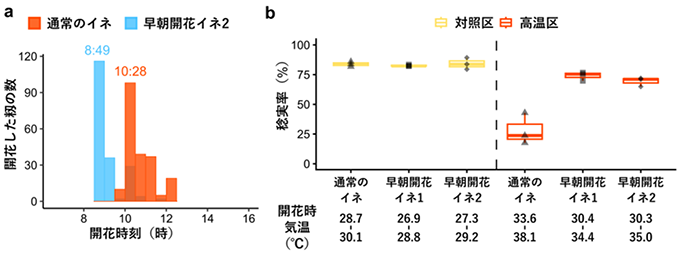
EMF3 遺伝子の塩基配列に変異を持つ日本の品種(「とよめき」)の突然変異系統群を用いて開花時刻を調査したところ、61番目のアミノ酸の変化の他に、563番目のアミノ酸がトレオニン(T)からイソロイシン(I)に変化しているイネも早朝開花性を示しました(早朝開花イネ2)(図4a )。さらに、午前6時の気温は28° C、その後、徐々に気温が上がり、正午には38° Cの高温に達する設定の人工気象室(高温区)で通常のイネと2種類の早朝開花イネを栽培したところ、通常のイネ品種では、野外条件(対照区)に比べて著しく稔実率(コメが実っている籾の割合)が低下する一方で、早朝開花イネでは高温になる前に開花するため、高温区でもそれほど稔実率が低下しないことが実証されました(図4b )。様々なイネ品種の遺伝子情報が載っているデータベース(TASUKE+;https://tasuke-wiki.dna.naro.go.jp/ )で検索する限りでは、世界のどのイネ品種も早朝開花イネのような61番目と563番目のアミノ酸の変化を持っておらず、本研究で開発した早朝開花イネは、既存のイネ品種が持っていない特徴を持つ非常にユニークなイネであることが分かりました。
図4 EMF3タンパク質の61番目(早朝開花イネ1)または563番目(早朝開花イネ2)のアミノ酸に変異を有する2種類の早朝開花イネを用いた高温不稔軽減の実証試験結果 ° C、その後、正午に38° Cになるように徐々に人工気象室内の気温を上昇させました。早朝開花イネ1と2は気温が35° Cに達する前に開花を終えることができたのに対し、通常のイネは35° Cに達した後にほとんどの籾が開花しました。その結果、通常のイネは高温区で稔実率の低下がみられたのに対し、早朝開花イネ1と2は高温区においても稔実率の低下はみられませんでした。野外の対照区では開花時刻が遅い通常のイネでも開花時の気温は30.1° Cで、高い稔実率である(高温不稔は発生していない)ことが分かります。
③ EMF3 遺伝子は世界の様々なイネの開花時刻を早めることができる
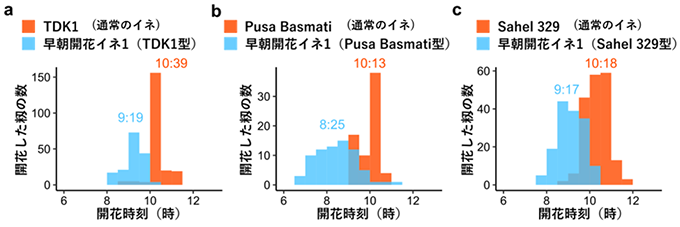
EMF3 遺伝子の変異は世界の様々なイネの開花時刻を早めることができることも分かりました。東南アジア(ラオス)で栽培されているTDK1、南アジアで栽培されているPusa Basmati、西アフリカで栽培されているSahel329という品種に、早朝開花イネ1で見られた61番目のアミノ酸が変化したEMF3 遺伝子をDNAマーカー6) を用いて交配により導入したところ、すべての型のイネで元の品種よりも開花時刻が早まりました(図5 )。
図5 EMF3 の61番目のアミノ酸の変化(早朝開花1)を導入した世界のイネの開花時刻。国際稲研究所(フィリピン)での結果
④ EMF3 遺伝子を通じ葯が開花時刻を制御する?
EMF3 遺伝子がどのように開花時刻を制御するのか、詳しいメカニズムの解明にも取り組みました。イネでは、鱗被(りんぴ)と呼ばれる花弁(かべん)7) に相当する器官に水分が流入することで鱗被が膨張し、物理的に花が開きます(図6a )。EMF3 遺伝子がイネの花のどの器官で働くのかを調査したところ、EMF3 遺伝子は鱗被ではなく、葯で発現していることが分かりました(図6b )。この結果はイネの開花時刻決定が鱗被の働きのみで説明できるほど単純ではなく、開花時刻の制御に関わるまだ知られていない重要な生物学的プロセスが存在することを示唆しています。EMF3 遺伝子の特定により、イネでは葯が開花時刻を制御する可能性が世界で初めて示されましたが、葯で発現するEMF3 遺伝子が、どのような仕組みで鱗被の膨張という開花のプロセスを促すのかは明らかになっておらず、今後の研究が必要です。
図6 鱗被の膨張によるイネの開花とEMF3 遺伝子の葯特異的な発現 EMF3 遺伝子の開花日の葯での特異的な発現。青色の部分はEMF3 遺伝子が発現している部分です(黒い枠の中)。左は籾の外側から見た写真、右は*印 のついた籾の中の写真です。(b)の白矢頭は鱗被の位置を示しています。
今後の予定・期待
本研究は、急速に進行する温暖化の中で問題となることが予想される高温不稔の解決策として、EMF3 遺伝子を利用した早朝開花イネの育成が有効である可能性を示しました。わずか1塩基の変化がイネの早朝開花性をもたらすことを明らかにしたことにより、国内外で精度の高いDNAマーカーの開発が進み、早朝開花イネの品種育成が加速すると期待されます。加えて、EMF3 遺伝子が鱗被ではなく葯で発現することを見出した本研究の知見は、葯を介した開花時刻の制御メカニズムの全容を科学的に解き明かす重要な手掛かりとなると考えられます。
用語の解説
登熟期
イネの穂が現れる時期を出穂期といい、出穂してから収穫までのコメが肥大し実る期間を登熟期という。登熟が完了すると収穫となる。[概要に戻る]
塩基配列
DNAやRNAなどの核酸において、アデニン(A)、チミン(T/RNAはU)、グアニン(G)、シトシン(C)の4種類の塩基が連なる「並び順」のこと。この配列が生物の遺伝情報(タンパク質の設計図)をコードしており、配列の順序が異なると、作られるタンパク質の性質や機能が変化する。[概要に戻る]
早朝開花性
通常より早い時間帯に開花する性質。イネは開花時に高温に対する感受性が最も高い一方で、開花が終了した1時間後に高温に遭遇した場合は高温不稔がほとんど発生しないことから、早朝開花性をイネに与えられれば、高温不稔を軽減できるという理論がSatake and Yoshida (1978)により提唱されていた。[概要に戻る]
葯(やく)
雄しべの先の、花粉が入った袋[開発の社会的背景に戻る]
柱頭(ちゅうとう)
めしべの頂部で花粉を受け取るところ[開発の社会的背景に戻る]
DNAマーカー
有用遺伝子のゲノム上の存在位置の目印となる塩基配列。目的の形質(この場合はEMF3 遺伝子に由来する早朝開花性)を持つ個体を効率的に選抜するため、例えば幼苗の段階で葉のDNAを分析し、EMF3 遺伝子のマーカーの有無で品種改良を行う。目的の遺伝子を持つ個体を迅速に見つけ出せるため、従来の育成法より品種開発の期間を大幅に短縮できる。[研究の内容・意義に戻る]
花弁(かべん)
一般では「花びら」のこと[研究の内容・意義に戻る]
発表論文
Takuma Ishizaki, Yoichi Hashida, Hideyuki Hirabayashi, Kazuhiro Sasaki, Hiroki Tokunaga, Eliza Vie M. Simon-Ada, Masataka Wakayama, Toshiyuki Takai, Hiroki Saito, Atsushi J. Nagano, Hitoshi Sakakibara, Mikiko Kojima, Yumiko Takebayashi, Sung-Ryul Kim, Ryo Matsushima, Michael J. Thomson, Kazuhiko Sugimoto, Ken-ichiro Hibara, Tsutomu Ishimaru. (2026) Rice EMF3 alleles adjust flower opening time to enhance the seed setting rate under high temperature stress. Plant Biotechnology Journal.
<著者情報>
農研機構
国際農林水産業研究センター
高崎健康福祉大学
吉備国際大学
国際稲研究所
龍谷大学(当時)/慶應義塾大学/名古屋大学
慶應義塾大学
名古屋大学大学院/理化学研究所
理化学研究所
岡山大学
研究担当者の声
イネの開花とコメについての講演
農研機構 中日本農業研究センター石丸 努
早朝開花イネの研究を始めてから19年間、毎年真夏の炎天下に大汗をかいて開花を観察してきました。人間はクーラーの利いた屋内で快適に過ごすことができますが、イネは屋内に避難することができません。「イネもせめて早朝に咲くことができれば、涼しくて快適だろうな」と思って実験していました。『早起きは三文の徳』という諺がありますが、進行著しい温暖化の中で早朝開花イネはまさに、その諺を体現したイネだと私はうれしく思います。