社会的背景
ニホンナシなどの落葉果樹では、一般的に収穫前年のうちに翌年の花芽が完成します。花芽は枝伸長が停止した後の初夏頃から形成が始まり、秋頃までに分化を終え、冬の休眠を経て翌年の春に開花します。近年、気候温暖化に伴い夏から秋が高温で推移する傾向が強まっています。このような気象条件では、高温により枝伸長が遅くまで続くことが観察されており、それに伴って花芽形成開始期も遅くなることから、冬の休眠までに花芽が完成できず、十分な花芽を確保できなくなる場合があります。
気候温暖化はさらに進展することが予測されています。今後も果実を安定して生産するためには、温暖化条件でも花芽を安定して確保できる技術の開発が必要になっています。
研究の経緯
多くの植物は日の長さや光の質の変化を感受して季節の移り変わりを認識し、花芽形成を開始する時期を決定しています。ところが、ニホンナシを含むバラ科ナシ亜科の果樹では、これまで花芽形成期の制御に光はほとんど関係せず、温度のみによって制御されていると考えられてきました。一方、最近の研究により、植物では、光と温度はシグナルの受容や伝達の経路が一部共有されていることが明らかにされています。そこで私達は、ニホンナシにおいて花芽形成を阻害する温度(高温)の影響を、光の処理によって打ち消すことができないかと考えました。
また、近年高性能化が進む発光ダイオード(LED)照明は、単波長の光を高いエネルギー効率で照射できる特長があり、農業分野での利用についても検討が進んでいます。そこで、ニホンナシの花芽形成における光の影響を解明するとともに、LED照明を利用した花芽形成制御技術を開発することにしました。
研究の内容・意義
1.ニホンナシの花芽形成を促進する光特性と、その制御機構を明らかにしました。
1)ガラス室内で日照時間を一日8時間に制限して栽培したニホンナシの樹に、4月上旬~8月上旬の夜間相当期に赤色光または遠赤色光を連続照射したところ、遠赤色光照射処理区では枝伸長停止期、花芽形成期ともに早まり、照射終了時には21~40%の花芽形成率となりました(図1、表1)。
2) 遠赤色光源として、遠赤色波長域を強化した蛍光灯1種と、ピーク波長の異なる遠赤色LED光源3種を供試したところ、花芽形成促進効果が認められたのはピーク波長730 nmのLED光源と遠赤色強化蛍光灯の2つでした(図2、2012年)。このことから、730 nm付近の波長の光が花芽形成促進に効果的であることが分かりました。
3) 光照射期間における茎頂の遺伝子発現解析から、花芽形成を抑制するTerminal Flower1(TFL1、ティーエフエル1)遺伝子2)のニホンナシ型(オルソログ3))であるPpTFL1-1a、1-2aの発現が、遠赤色光照射により抑制されることが分かりました(図3)。
2.夜間遠赤色光照射は猛暑の翌年の花芽数減少を緩和することを示しました。
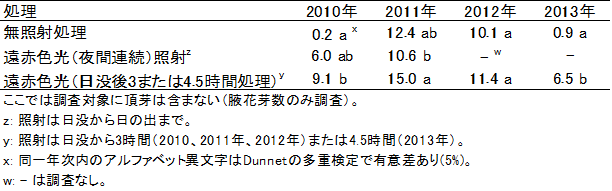
ほ場植栽のニホンナシ「幸水」では、開花前(4月)から落葉期(10月)までの夜間に遠赤色光を照射すると、猛暑で花芽形成数が少ない年(2010年、2013年)でも花芽形成の減少が抑えられ、ほぼ例年並みの花芽数が得られました。しかし、花芽形成数が例年並みの年(2011年、2012年)では、更なる促進効果は認められませんでした(表2)。処理時間については、日没から日没後3または4.5時間の短い照射でも、夜間連続照射と同等以上の高い効果が認められました(表2)。
今後の予定・期待
バラ科果樹のニホンナシにおいて花芽形成への光の関与が初めて示されたことから、今後は開花や休眠など、これまで光が無関係と考えられていた現象に関しても光の役割を再検討する必要があります。また、リンゴやセイヨウナシなどのニホンナシ以外のナシ亜科果樹についても光による生育制御の可能性が考えられます。
私達がほ場で照射した光強度は枝発生面で受光量1~2 mmol s-1 m-2でした。現在入手可能な遠赤色LED光源は出力が弱いため、試験では2.4個/m2の密度で配置した5WのLED電球を枝発生面1.5mの距離から照射しており、この光量を栽培現場で実現するのはコスト的に難しいと考えられます。今後は、光照射による花芽形成の効率化に向けて、果樹の光に対する反応をより詳細に検討し、光照射時期の絞り込みや照射強度の低減化について検討を進める必要があります。また、LED光源は、長寿命・省エネルギーの特性をもち、世界的にその高性能化が図られている状況にあります。本研究の成果により果樹生産に対する遠赤色光領域のLED光源の有効性が明らかとなったことで、今後、効率的な照射方法と、より安価で高性能なLED光源が開発され、近い将来「電照果樹栽培」が実現するかもしれません。
用語の解説
1) 遠赤色光
光の色は波長で決まります。光のうち、人間が感知できる範囲は概ね380~780 nmの範囲とされており、その範囲を「可視光領域」と呼びます。可視光より短い波長域にX線や紫外線、可視光より長い波長域に赤外線や電波があります。
波長400~800 nmの光は、青色部(B 400~500 nm)、緑色部(G 500~600 nm)、赤色部(R 600~700 nm)、遠赤色部(FR 700~800 nm)と区分されます(※ 波長領域と光色との対比は調査や定義により異なる場合があります)。遠赤色部に区分される光のことを「遠赤色光」と言います。
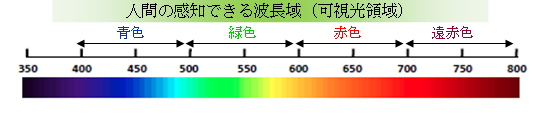
(農研機構花き研究所「キク電照栽培用光源選定・導入のてびき(2013年)」を元に作成)
2) Terminal Flower 1(TFL1)遺伝子
植物の花芽形成力を弱めたり阻害したりする機能を持つ遺伝子で、花成ホルモンの本体であり植物の花芽形成を誘起するFlowering Locus T(FT)遺伝子の働きを抑えます。シロイヌナズナ(アラビドプシス)において発見された遺伝子ですが、リンゴでもリンゴ型のTerminal Flower 1遺伝子の発現を抑えた変異体で、開花が早まることが確認されています。
3) オルソログ
異なる種の遺伝子やタンパク質で、その塩基配列やアミノ酸配列が非常によく似ているため、同一の祖先遺伝子に由来し、類似の機能を持つと推定されるもの。TFL1遺伝子は、実験植物のシロイヌナズナ(アラビドプシス)で構造と機能が明らかにされており、PpTFL1-1a、1-2aはこれと塩基配列が類似しているニホンナシの遺伝子です。
本成果の論文発表
論題:Effect of extending the photoperiod with low-intensity red or far-red light on the timing of shoot elongation and flower-bud formation of 1-year-old Japanese pear (Pyrus pyrifolia).
(赤色または遠赤色光による日長の延長が1年生ニホンナシ苗の枝伸長停止期および花芽形成期に及ぼす影響)
掲載紙: Tree Physiology 34, 534–546 (2014)
著者: 伊東明子、齋藤隆徳、西島隆明、森口卓哉
図表
A:無照射樹

B:遠赤色光照射樹

図1
日照時間を8時間に制限したガラス室内で夜間相当期に4ヶ月の光照射処理後、高温・長日条件で3週間管理し、萌芽させた茎頂の様子。
A:無照射(左)の樹では茎頂に花芽が形成されていないため枝葉が伸長しているが、B:遠赤色光照射樹(右)では花蕾が出現している。
表1 ガラス室内での夜間相当期の4ヶ月の光
照射が茎頂の花芽形成率(%)に及ぼす影響
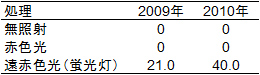
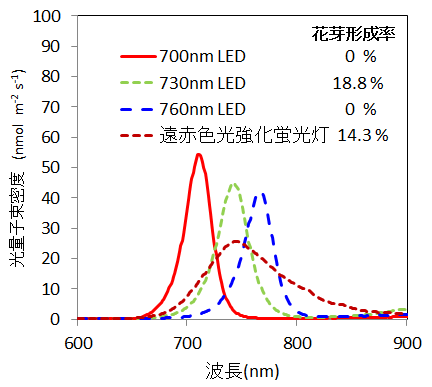
図2 光照射処理に使用した光源の光波長と、
照射4ヶ月後の茎頂花芽形成率
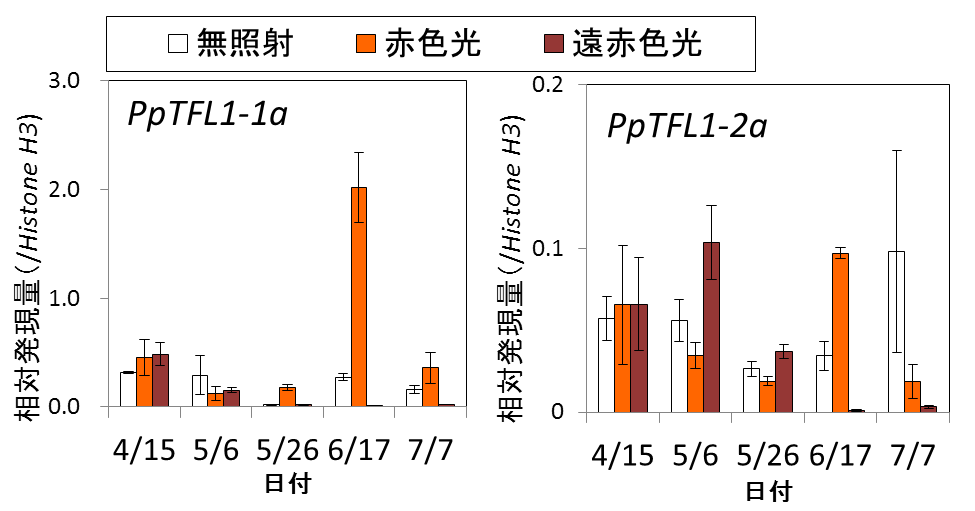
図3 ガラス室内での夜間相当期の4ヶ月間の光照射が茎頂の遺伝子発現に及ぼす影響(図中の縦棒はサンプル間のばらつきの範囲を示す)。遠赤色光を照射した茎頂では、花芽形成時期(6月以降)になるとこれらの遺伝子発現が抑えられ、花芽が形成される。
表2 ほ場植栽のニホンナシ「幸水」樹に対する夜間の遠赤色光照射処理が新梢あたり花芽数に及ぼす影響(2010~2013年)