開発の社会的背景
イネやムギなど、日本で栽培されている作物の多くは、弥生時代以降に大陸から伝播したもので、これまで、アズキ(図1 )もその一つだと考えられていました。一般に、DNA配列の多様性は起源地で最も高く、そこから離れるにしたがって低下します。DNAマーカーを用いた栽培アズキの遺伝的多様性解析では、中国南部が最も多様性が高くなっていたことから、中国起源説が支持されていました。一方、日本では近年、縄文時代の遺跡から多数のアズキ種子が発掘されており、縄文時代後期の遺跡から発掘されたアズキの種子は、同時代の中国や韓国の遺跡から発掘されたものより大型でした。栽培化に伴って種子は大型化する傾向があることから、アズキの日本起源説が提唱されはじめました。しかし、種子の大きさが環境条件に左右されやすいことは、考古学的な証拠に基づく日本起源説におけるウィークポイントとなっていました。
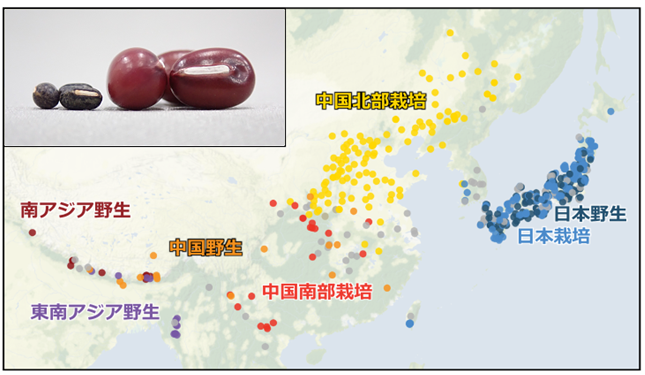
図1. 解析に使用した栽培アズキ(栽培)およびヤブツルアズキ(野生)の採取地遺伝的構造による7グループごとに色分けをしている。灰色は、雑種など遺伝的な分類が曖昧なもの。栽培アズキは東アジアだけでなく、ネパール・ブータンにまで広がっている。左上はヤブツルアズキ(左)と栽培アズキ(右)の種子。
研究の経緯
農研機構と台湾大学の国際共同研究グループは、日本や中国を含めたアジア各地から収集された栽培アズキおよびヤブツルアズキ、計693系統の全ゲノム配列を解析しました。その結果は、栽培アズキにおける核ゲノムの多様性は中国南部で最も高く、これまで通り中国起源説を支持するものでした。
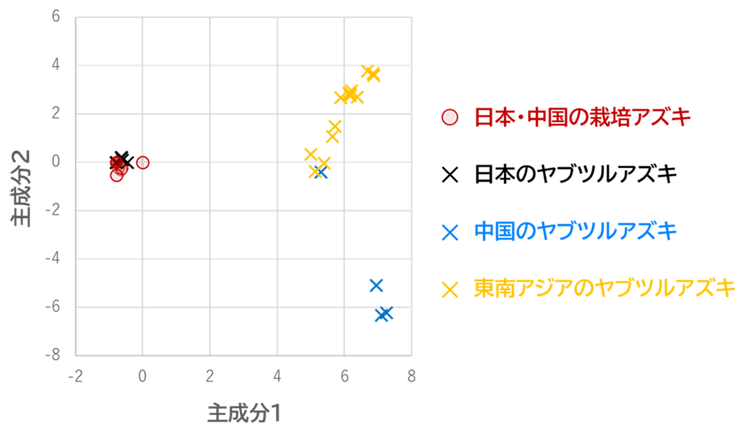
一方、葉緑体のゲノムは全ての栽培アズキが日本の野生種であるヤブツルアズキと同一の型を示す、という前述の結果に相反するものとなりました(図2 )。栽培アズキと野生種が互いに受粉して雑種を形成しても、葉緑体は母性遺伝するため必ず種子親(母親)の葉緑体ゲノムを受け継ぎます。ヒトがアズキを栽培するようになってからは、「外部のヤブツルアズキの花粉が畑のアズキに受粉してできた雑種種子」が収穫されることはあっても、その逆は稀です。すなわち、現在の栽培アズキがもつ葉緑体ゲノムは、最初期の栽培アズキからずっと受け継がれてきたと考えられます。したがって、仮に中国起源説が正しい場合、栽培アズキの葉緑体ゲノムは日本のヤブツルアズキよりも中国のヤブツルアズキに近くなるはずです。しかし、中国の栽培アズキを含む全ての栽培アズキが例外なく日本のヤブツルアズキと同じ葉緑体ゲノムを有すると判明しました。このことは、栽培アズキの日本起源説を支持する強力な遺伝学的な証拠となります。
図2. 栽培アズキおよびヤブツルアズキの葉緑体ゲノム配列に基づく主成分分析系統間の近縁関係を示したもので、近縁なもの同士ほどグラフ上の位置が近くなる。
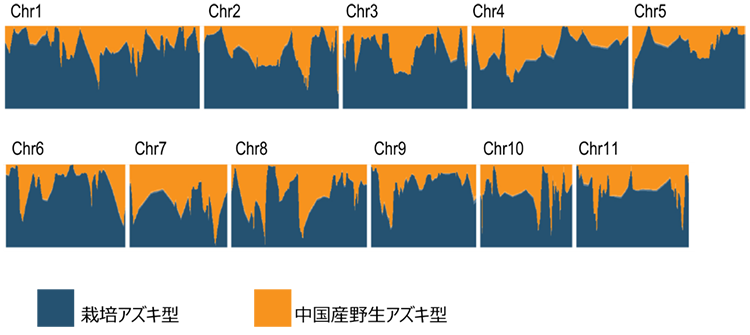
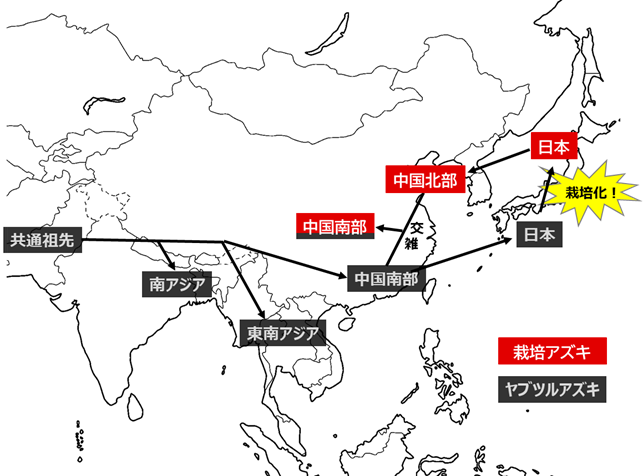
次に、栽培アズキにおける核ゲノムの多様性が中国南部で最も高い原因を明らかにするために、研究グループは中国南部の栽培アズキのゲノムを、日本の栽培アズキのゲノム及び中国のヤブツルアズキのゲノムと詳細に比較することにしました。その結果、中国南部の栽培アズキの核ゲノムは、部分的に同地域のヤブツルアズキのものに置き換わっていることが明らかとなりました(図3 )。すなわち、中国南部の栽培アズキは、日本から持ち込まれた栽培アズキに現地のヤブツルアズキの花粉が交雑して成立したものであり、中国南部における栽培アズキの高い多様性は、現地のヤブツルアズキに由来することが明らかとなりました。この結果は、アズキの日本起源説を支持する結果です。(図4 )。
図3. 中国の栽培アズキの核ゲノム構成Chr1からChr11は染色体1番から11番を指す。
図4. ゲノム解析結果から推察される、ヤブツルアズキおよび栽培アズキの伝播経路・成立過程日本から中国北部を経て持ち込まれた栽培アズキに、現地のヤブツルアズキが交雑することで、多様性が増加した栽培アズキ集団が成立した。
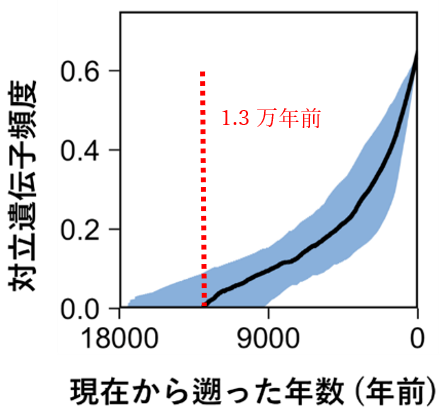
さらに、アズキの栽培化がいつ始まったのかを、栽培アズキとヤブツルアズキを区別する特徴として重要な「種皮の色」の遺伝子ANR1を用いて解析しました。ヤブツルアズキでは黒味をおびている種皮の色が、栽培アズキではANR1遺伝子に起きた突然変異によって、赤くなります (図1 )。また、この変異は種皮の透水性3) にも影響するため、自然界での生き残りには不利ですが、播種後の発芽の斉一性・調理適性・人の赤い色への嗜好性などから人為的な選抜の対象になります。したがって、栽培と選抜を繰り返すうちに、栽培アズキの集団では変異型の赤いアズキばかりになったと考えられます。多数のサンプルのゲノム情報をもとに、栽培アズキでの変異型遺伝子ANR1の頻度(対立遺伝子頻度)の変化を推定したところ、約1万世代前から増加し始めたことが示唆されました(図5 )。上述の通り、自然界で不利な変異型ANR1の頻度が増加したのは、人が赤いアズキを選抜していたことを裏付けています。アズキは一年生の植物であることから、人類によるアズキの栽培は、約1万年前には始まっていたと考えられます。日本にイネが伝来して稲作が始まったのは約3,000年前とされていますが、日本列島におけるアズキの栽培はそれよりも遥かに以前から始まっていた可能性が示されました。
図5. 栽培アズキにおける変異型ANR1遺伝子の相対頻度の推移青で示した領域は95%信頼区間4) 。赤いアズキの頻度は約1.3万年前(信頼区間 : 1.8万年前から0.9万年前)から増加が始まり、現在には赤いアズキが大半を占めるに至った。
研究の内容・意義
本研究により、栽培アズキの日本起源説を支持する結果が得られました。また、アズキの栽培化が少なくとも1万年前から始まった可能性があることは、縄文時代後期の日本でアズキが栽培されていたとする考古学的な仮説を支持しています。
今後の予定・期待
作物の起源地や進化を明らかにすることは、現在、利用されている遺伝子や未利用の遺伝子を明らかにし育種に活用できる新たな知見をもたらす研究です。今回のアズキの解析事例を他の作物にも応用することによって、未利用遺伝子の活用がより一層進むものと期待されます。
用語の解説
栽培化
野生植物が人為的な選抜によって作物となること。野生植物は自然環境での生存・繁殖に有利な形質を有しますが、栽培化された植物は人類による栽培・収穫・利用に適した形質をもつものが優先的に選抜された結果、しばしば元の野生植物とは全く異なる姿形となります。大型化は、栽培化の際に選抜されやすい重要な形質の一つです。
[ポイントへ戻る]
母性遺伝
花粉親(雄)と種子親(雌)が交雑してできた子の核ゲノムには、両親のゲノムが50%ずつ受け継がれます。一方、葉緑体やミトコンドリアは、花粉親由来のものは分解され、種子親由来のものだけが受け継がれます。
[概要へ戻る]
種皮の透水性
ヤブツルアズキの種皮は全く水を通さず、種皮が分解されるか機械的に傷つけられるかしない限りは吸水せず、発芽しません。この性質が残っていると、畑に種を撒いて水をやっても芽が出てこないため、栽培には不向きです。栽培アズキの種皮はこの性質が失われており、水をやれば、種子は容易に吸水し発芽します。栽培アズキでは種皮の色の変化と透水性の変化には強い相関があるため、赤いアズキが選抜された理由は見た目だけではなく、畑に蒔いたときに発芽しやすいものが選抜されたことの影響もあったと考えられます。
[研究の経緯へ戻る]
95%信頼区間
このような研究では、実験で得られた結果に基づいて、もともとの現象ではどのくらいの値になるかを統計学的手法で推定します。このときに、真の値がその範囲に収まる確率が95%の区間を指します。
[研究の経緯(図5)へ戻る]
発表論文
Chih-Cheng Chien, Takashi Seiko, Chiaki Muto, Hirotaka Ariga, Yen-Chiao Wang, Chuan-Hsin Chan, Hiroaki Sakai, Ken Naito, Cheng-Ruei Lee (2025) A single domestication origin of the adzuki bean and the evolution of domestication genes. https://www.science.org/doi/10.1126/science.ads2871