開発の社会的背景と研究の経緯
マメ科植物は根粒菌と呼ばれる土壌細菌と共生し、根に根粒を形成します。根粒菌は根粒の中で空気中の窒素をアンモニアに変換する窒素固定を行い、固定した窒素をマメ科植物に供給することで、マメ科植物の生長に大きく貢献します。マメ科植物は、この共生窒素固定の能力を活用することで、窒素肥料に依存しない栽培が可能になると期待されています。一方で、土壌中に存在する根粒菌の中には、マメ科植物に住み着き根粒を形成するにも関わらず、窒素固定を行えない不良根粒菌が複数存在しています。現在の農業生産において、それらの不良根粒菌が、土壌中や人工的に接種した優良根粒菌の共生を妨げる事例が知られており、期待した共生窒素固定の効果が得られないことが問題となっています。apn1 変異体)と、その変異体において窒素固定を行えないミヤコグサ根粒菌を用い、窒素固定がうまく行かない原因を明らかにすることで、窒素固定活性を発現するメカニズムを解明することを目指しました。
研究の内容・意義
窒素固定活性の低下を引き起こす根粒菌遺伝子DCA1 の同定 研究チームは、根粒菌株に応じて異なる窒素固定活性を示すミヤコグサの突然変異体(
apn1 変異体)において、窒素固定できない根粒菌(TONO菌)の遺伝子を破壊したTONO菌変異株の中から、
apn1 変異体でも窒素固定できるようになったTONO菌変異株を選抜しました(
図1 )。この変異株では、
グラム陰性細菌3) に広く存在する
オートトランスポーター4) と呼ばれるタンパク質分泌装置をコードする遺伝子が変異し、その機能を失っていました。研究チームはこの根粒菌の遺伝子を
DCA1 (Determinant of nitrogen fixation Compatibility of APN1)と名付けました。
DCA1は根粒菌の細胞膜を貫通する様な構造を持ち、自らのタンパク質の一部を切り取り、菌体外に分泌することがわかりました。DCA1に類似した構造を持つタンパク質は、ミヤコグサ根粒菌以外の根粒菌や他のグラム陰性細菌、さらにはシアノバクテリアにも存在していますが、その機能は明らかになっていませんでした。その中で研究チームはDCA1が植物と微生物共生に関与することを初めて明らかにしました。
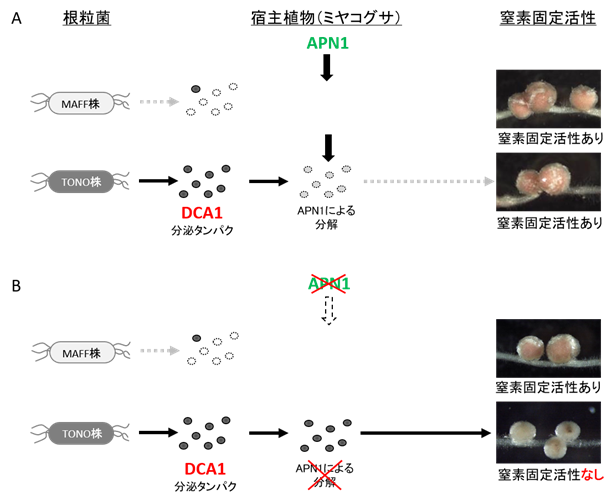
マメ科植物の共生戦略 ミヤコグサのAPN1は
アスパラギン酸ペプチダーゼ5) と呼ばれるタンパク質分解酵素で、根粒で働くことが分かっています。ミヤコグサのAPN1と根粒菌のDCA1の組換えタンパク質を作製し、両者を反応させたところ、APN1はDCA1を分解する活性を持つことがわかりました。APN1が機能する野生型ミヤコグサの根粒では、根粒菌のDCA1が働いても正常な窒素固定が起こりますが、APN1が壊れている
apn1 変異体の根粒では、DCA1が分解されないため窒素固定活性が低下します(
図2 )。このことから、窒素固定を低下させる根粒菌のDCA1の効果を、宿主植物のAPN1が打ち消すことによって、正常な窒素固定活性が発現できるように宿主植物が根粒菌の窒素固定機能を制御していることがわかりました。
今後の予定・期待
APN1によるDCA1分解機構の発見は、宿主植物に共生しながら、窒素固定を行わない根粒菌と、そのような根粒菌を取り締まる、宿主植物との間の複雑な関係性を明らかにしました。
用語の解説
根粒菌
ダイズなどのマメ科植物の根に感染し、根粒と呼ばれるこぶ状の器官を形成する土壌細菌。根粒菌は根粒の中で空気中の窒素をアンモニアに変換(窒素固定)し、宿主植物に供給します。宿主植物はその見返りとして光合成産物を根粒菌に供給します。マメ科植物と根粒菌の共生により窒素固定反応が起きる現象を、ここでは共生窒素固定と呼んでいます。
ミヤコグサ(学名:Lotus japonicus )
日本を含めアジアの温帯地域に自生する多年生のマメ科植物。ダイズなどの農業上重要なマメ科作物に比べ、ゲノムサイズが小さい、形質転換が容易、植物体のサイズが小さく世代時間が短い、などの分子遺伝学的解析に適した性質を持つことから、マメ科のモデル植物に位置付けられ、根粒菌との共生に関する研究をはじめ様々な分野に利用されています。
グラム陰性細菌
すべての細菌はグラム陽性菌とグラム陰性菌に分類されます。デンマークの学者ハンス・グラムが開発した細菌の染色法で染まるものをグラム陽性細菌、染まらないものをグラム陰性細菌と呼びます。両者は細胞壁の構造に大きな違いがあり、グラム陰性菌は細胞壁の外側に外膜を持ち、外膜から特定のタンパク質や核酸を分泌する装置を持っています。
オートトランスポーター
グラム陰性細菌が持つタンパク質分泌装置の1つ。別名5型タンパク質分泌装置。グラム陰性細菌のタンパク質分泌装置は構成するタンパク質や分泌様式から1型から6型に分類されます。オートトランスポーターは細胞膜に孔を形成し、自身のタンパク質の一部を細胞外に分泌あるいは細菌表面に露出させます。
アスパラギン酸ペプチダーゼ
ペプチダーゼはタンパク質やポリペプチドを加水分解する酵素の総称。触媒機構によっていくつかのグループに分類されます。アスパラギン酸ペプチダーゼは活性中心にアスパラギン酸残基を持ち、主に酸性条件で働くペプチダーゼ(代表例:ペプシン)。
発表論文
The rhizobial autotransporter determines the symbiotic nitrogen fixation activity of Lotus japonicus in a host-specific manner
Loss-of-function of ASPARTIC PEPTIDASE NODULE-INDUCED 1 (APN1) in Lotus japonicus restricts efficient nitrogen-fixing symbiosis with specific Mesorhizobium loti strains
参考図
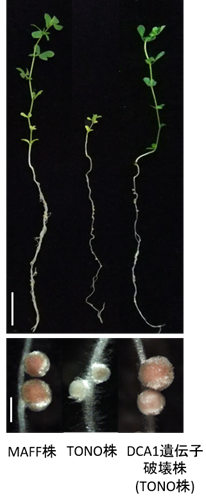
図1 根粒菌株に応じて異なる窒素固定活性を示すミヤコグサのapn1 変異体 apn1 変異体の様子。スケールバー:2cm。apn1 変異体の根粒。スケールバー:1mm。 apn1 変異体はミヤコグサ根粒菌MAFF株を接種した場合には、窒素固定活性のある根粒を形成し、窒素栄養を与えない条件でも地上部がよく成長する(左写真)のに対し、ミヤコグサ根粒菌TONO株を接種した場合には、窒素固定活性の低い根粒を着生し、地上部がほとんど成長しません(中央写真)。一方、DCA1 遺伝子を破壊したTONO株はapn1 変異体において窒素固定活性のある根粒を形成できます(右写真)。図2 ミヤコグサと根粒菌の共生窒素固定活性を決定するメカニズム DCA1 )を有しますが、TONO株はMAFF株に比べてDCA1を多量に合成します。A:APN1が機能する野生型ミヤコグサの場合。MAFF株、TONO株のDCA1から分泌されるタンパクがAPN1によって分解されることで、窒素固定活性の低下が阻止され、窒素固定活性を持つピンク色の根粒ができます。B:APN1が機能しないapn1 変異体の場合。 MAFF株はDCA1分泌タンパクの量が少ないため、APN1によるタンパク分解がなくても窒素固定活性は維持されます。一方、TONO株が感染した根粒では、TONO株が合成する多量のDCA1分泌タンパクが分解されないため窒素固定活性が低下し、窒素固定活性のない根粒が形成されます。