研究の社会的背景
世界におよそ3億haある灌漑農地の約半分は塩類集積による塩害の影響を受けており、塩害に極めて弱いイネやダイズなど主要な作物の栽培が困難になっています。また、乾燥地での灌漑農業が拡大した結果、湖沼や地下水などの淡水資源が急速に枯渇に向かっています。このような状況に対して、塩水でも栽培可能な塩害に強い作物の開発が求められており、塩害に強い植物がもつ耐塩性機構を明らかにすることが不可欠となっています。
研究の経緯
農研機構では、多様な遺伝資源1) をジーンバンク2) 事業で収集・保存し、有用な特徴を調べています。特にアズキの近縁種は多様性の宝庫であり、厳しい環境に適応した種や系統が多数存在します。私たちは、アズキの遺伝資源が種ごとに独自の耐塩性機構を有すること(Noda et al., 2022)や、ヒナアズキがデンプンを利用した特殊な耐塩性機構を有すること(Noda et al., 2023)を明らかにしてきました。
本研究では、アズキ近縁種の中でも最も塩害に強く、塩水に浸かっても、植物体内へのナトリウムの取り込みを強く制限できるハマササゲを対象としました(図1 )。葉や茎へのナトリウム流入を防ぐ耐塩性植物は多く知られていますが、根にも蓄積しない植物は異例です。本研究グループは、QSTと筑波大学が有する元素可視化技術、東京大学が有する元素定量技術、そして、農研機構のゲノム解読・解析技術を用いて、ハマササゲがもつ耐塩性機構の真相究明に取り組みました。
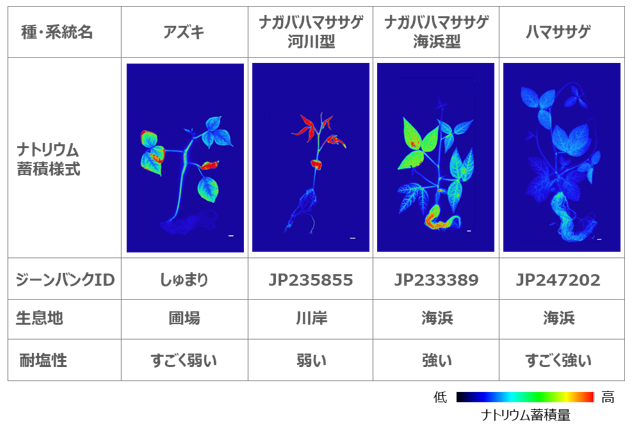
図1. アズキ、ナガバハマササゲ(河川型・海浜型)およびハマササゲのナトリウム蓄積様式および特性情報
【ハマササゲは高いナトリウム排出能力をもち、ナトリウム排出には日周性がある】
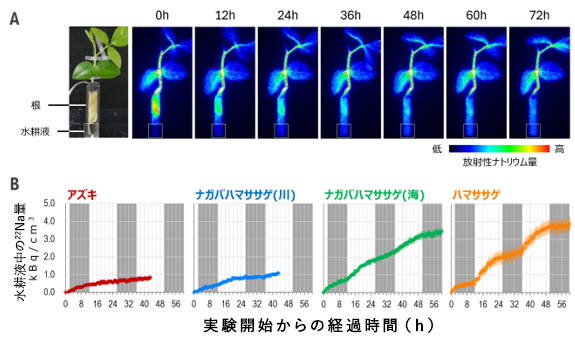
ハマササゲの耐塩性を明らかにするため、QSTが開発した植物用のPositron-Emitting Tracer Imaging System (PETIS3) : ポジトロンイメージング装置)と放射性ナトリウムを使い、植物の各器官や水耕培地におけるナトリウムの濃度変化をリアルタイムで追跡しました。その結果、ハマササゲの根に一旦ナトリウムが取り込まれ、その後根から排出されることが明らかとなりました(図2A )。根からのナトリウム排出量は昼間に増加し、夜間に減少していました。根からの吸水量は昼間に増加して夜間には減少することから、根へのナトリウムの流入も吸水の多い昼間に増加します。ナトリウム流入の少ない夜間にナトリウムの排出を停止することは、余計なエネルギー消費を抑える効果があると考えられます。
次に、根からのナトリウム排出やその日周性がハマササゲに特有の現象なのかを明らかにするため、ナガバハマササゲやアズキなど、耐塩性レベルの異なる3系統(図1 を参照)を加え、解析しました。その結果、根からのナトリウム排出量はアズキが最も少なく、ハマササゲで最も多くなり、耐塩性レベルと一致した結果となりました(図2B )。一方、排出の日周性に関しては、ハマササゲとナガバハマササゲには見られましたが、アズキでは観察されませんでした。
これらの結果から、ハマササゲは高いナトリウム排出能力を獲得したこと、また排出の日周性は、ハマササゲとナガバハマササゲの共通祖先において獲得されたものと考えられました。
図2. PETISによるナトリウム排出の解析
A. ハマササゲのPETIS解析。最初は根に大量に含まれている放射性ナトリウムが時間経過に伴い根から水耕液へと移動している様子が伺える。B. 水耕液中の放射性ナトリウム量の経時変化。塩害に強い系統ほどナトリウム排出速度が大きく、ハマササゲおよびナガバハマササゲは昼(白)には夜(灰色)よりも排出速度が速い日周性を示している。
【ハマササゲのナトリウム排出の特性は2遺伝子の発現パターンで説明できる】
ハマササゲのナトリウム排出に関わる遺伝子の同定のため、ハマササゲを含めた4系統(図1 )を塩ストレス下で栽培し、昼と夜にサンプリングした根からRNAを抽出して全遺伝子の発現解析を実施しました。
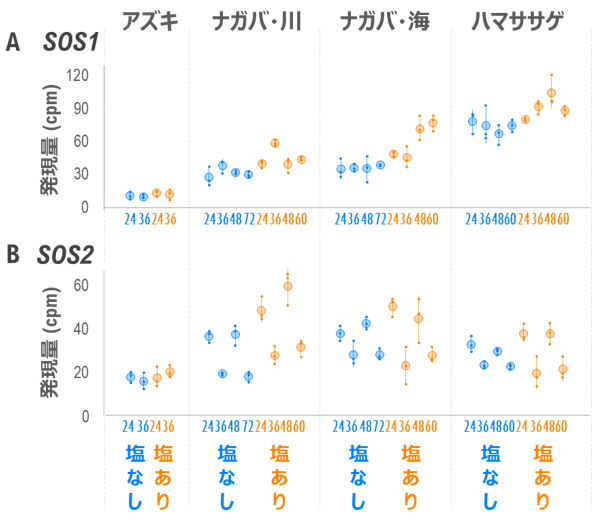
その結果、細胞内に流入したナトリウムを排出するナトリウム輸送体をコードするSOS1 遺伝子4) 図3A )。これに対し、塩害に弱いアズキでは塩ストレス下でもSOS1 遺伝子の発現は低く、ナガバハマササゲでも塩ストレスに応じて発現は上昇するものの、ハマササゲには及びませんでした。
また、SOS1 遺伝子の活性化に必要なSOS2 遺伝子5) 図3B )。以上から、ハマササゲを含む4系統に見られるナトリウム排出の特徴が、SOS1 およびSOS2 の2遺伝子の発現様式で説明できることが明らかとなりました。ナトリウム排出能力の最大値はSOS1 遺伝子の発現量で決まり、またナトリウム排出の日周性の有無はSOS2 遺伝子の発現様式で決まると考えられました。
図3. SOS1 およびSOS2 遺伝子の転写量の経時変化と系統間比較
横軸上の数値は塩ストレス開始(ゼロ日目の午前11時)からの経過時間を表し、24および48時間後のサンプルは昼に、36および60時間後のサンプルは夜に採取した。ナガバ・川(耐塩性低い)およびナガバ・海(耐塩性高い)はそれぞれナガバハマササゲの河川型と海浜型。
【ハマササゲの根の内皮には発達したリグニン層が形成される】
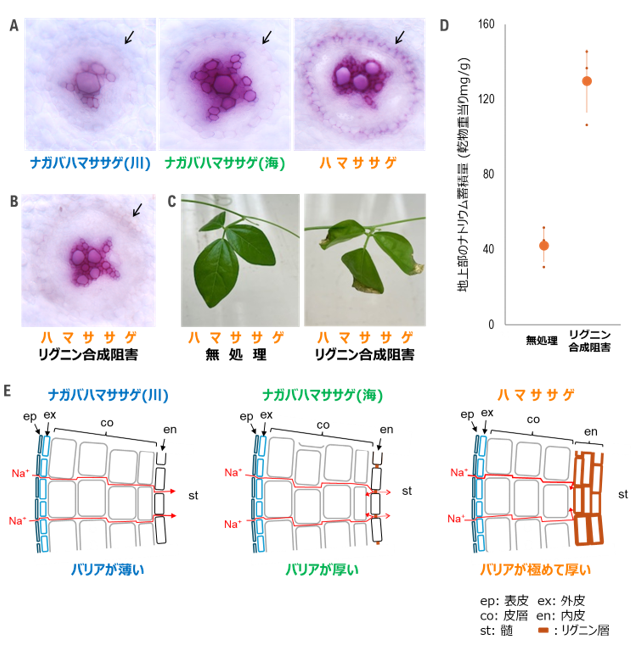
さらに、ハマササゲの耐塩性機構に関わる他の遺伝子を見つけるために、研究グループは全遺伝子発現解析結果から、ハマササゲで高い発現量を示す遺伝子群の機能を解析し、これらの遺伝子群には根のカスパリー線の形成に関わる遺伝子群が多いことを明らかにしました。カスパリー線とは、内皮の細胞間隙を埋めるようにリグニンなどのイオンを透しにくい物質が蓄積した構造をいいます(図4A、4E )。カスパリー線の存在により、外皮や皮層を通して水と一緒に流れ込んできた物質が、いったんは内皮細胞の内部を通り抜けないと道管にたどり着けないようになります。この内皮細胞には様々な輸送体があり、これらが必要な物資のみを選択的に内側方向へ輸送し、不要な物質は外側方向へ排出するフィルターのような役割を果たしています。したがって、ハマササゲにおいてカスパリー線に関わる遺伝子群の発現量が高いことは、ナトリウムを通さないバリアが発達している可能性を示すと考えられました。
そこで、塩ストレス下で栽培したハマササゲの根のリグニンを染色して顕微鏡で観察しました。一般の植物では1層の内皮細胞の細胞間隙にのみカスパリー線が形成されるのに対し、ハマササゲでは複数の細胞層にわたって多量のリグニンが蓄積している様子が観察されました(図4A )。
図4. 内皮に形成されるリグニン層の比較と、リグニン合成阻害がおよぼす塩害耐性への効果
A. 根の横断切片の顕微鏡写真。赤く染色されているのがリグニン。矢印は根の内皮の位置を示す。ハマササゲは内皮のリグニンがはっきりと赤く見える。B. リグニン合成阻害剤を投与されたハマササゲの根。矢印は内皮の位置を示す。C. 塩ストレス下で栽培したハマササゲにリグニン合成阻害剤を投与した様子。葉が色あせて傷害を生じている。D. 植物体の地上部に蓄積されたナトリウム量。Cの植物体から地上部を採取して測定した。E. 根におけるNa+ の輸送経路とおよびNa+ 流入抑制におおいてカスパリー線(内皮リグニン層)が果たす役割。カスパリー線のリグニンがハマササゲでは極めて厚く、Na+ が入りにくい。
【リグニンの合成阻害は、ハマササゲの耐塩性を弱体化させる】
リグニンの効果を明らかにするために、リグニン合成の阻害剤を植物に投与する実験を行いました。その結果、内皮のリグニン層は全く観察されなくなり(図4B )、この状態のハマササゲを塩ストレス下で栽培すると、葉に顕著な傷害が現れました(図4C )。塩ストレス下での植物体内のナトリウム蓄積量は、リグニン合成阻害剤を投与すると、地上部に流入するナトリウムの量が大幅に上昇しました(図4D )。したがって、ハマササゲは根の内皮に分厚いリグニン層を形成することで、ナトリウムを透さない強力なバリアを形成していることが明らかとなりました。
研究の内容・意義
【ハマササゲがもつ"最強"の耐塩性機構】
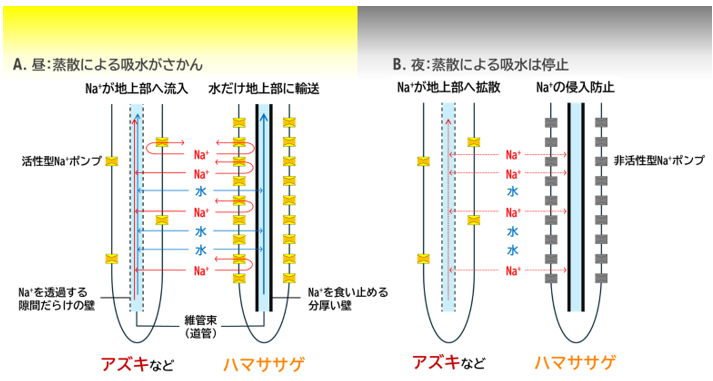
以上の結果から、今回明らかになったハマササゲの耐塩性機構は以下のようなものだと考えられます(図5 )。
根の内皮に分厚いバリアを形成し、ナトリウムが道管に流入することを防ぎます。バリアに食い止められたナトリウムは根に蓄積しますが、ハマササゲはナトリウム輸送体のSOS1 遺伝子を多量に発現しているため、根に留まっているナトリウムを外部に排出できます。これにより、ハマササゲは地上部だけでなく根のナトリウム蓄積量も低く保つことができると考えられます(図5A )。
夜間にSOS1 が非活性型になるのも、バリアのおかげだと考えられます。夜間には葉の気孔が閉じるため、根からの水の吸収もほぼ停止します。それでも、もし内皮のバリアがなければ、濃度勾配にしたがい根から地上部へと道管を通してナトリウムが拡散していきますが、バリアがあるハマササゲではそれは起きません。したがって、夜間のナトリウム排出を停止し、余計なエネルギーの消費を抑えることができると考えられます(図5B )。
図5. ハマササゲ耐塩性機構の概略図
A. 吸水が盛んな昼、ナトリウムは水と一緒に根に取り込まれ、アズキなどの塩害に弱い植物では道管まで浸透して地上部に輸送されるのに対し、ハマササゲでは内皮のバリアがあるためナトリウムは道管へは侵入できず、多くの活性型ポンプによって根から排出される。B. 吸水が停止する夜、アズキなど塩害に弱い植物ではナトリウムは道管を通して地上部まで拡散するが、ハマササゲでは道管への浸透は内皮によって防がれるため、ナトリウム排出ポンプが非活性型であっても地上部へのナトリウム拡散が起きない。
【ハマササゲ耐塩性機構の応用可能性】
今回の発見の重要な部分は、ハマササゲの耐塩性機構を構成する要素のいずれもが多くの植物に共通して存在するということです。イネやダイズなどの作物でも、もともと持っているこうした遺伝子の発現を増幅すればハマササゲの耐塩性機構を再現できる可能性があります。
今後の予定・期待
本研究によりナトリウム排出に関わる遺伝子は明らかとなりましたが、バリア形成に必要な遺伝子はまだ明らかになっていません。今後、この原因遺伝子を同定する予定です。また、他のアズキ近縁種がもつ別の耐塩性機構も明らかにし、耐塩性育種に繋げたいと考えています。
用語の解説
遺伝資源
遺伝の機能的な単位を有する植物、動物、微生物に由来する素材であって、顕在的または潜在的な価値を有するもの。
[研究の経緯へ戻る]
ジーンバンク
生物多様性の保全のほか、新品種や医薬品の開発等に活用するため、植物、動物、微生物の遺伝資源を収集し、人工的に管理することで、保存、配布する仕組みまたは施設。農研機構農業生物資源ジーンバンク : https://www.gene.affrc.go.jp/
[研究の経緯へ戻る]
PETIS(Positron-Emitting Tracer Imaging System)
ポジトロン放出核種をトレーサーとして、一対の検出器で消滅γ線を検出することにより、生きた植物体の中での物質をリアルタイムでマッピングすることにできる装置。生きた植物体の中の物質の動きが詳細にわかる。
[研究の経緯へ戻る]
SOS1 遺伝子
細胞内に流入したナトリウムイオンを細胞外へと輸送するナトリウム輸送体をコードした遺伝子。ナトリウムイオンを一つ排出するのと引き換えに、水素イオンを細胞内に取り込むアンチポーターである。
[研究の経緯へ戻る]
SOS2 遺伝子
SOS1タンパク質をリン酸化により活性化するリン酸化酵素をコードした遺伝子。SOS2タンパク質が存在しないとSOS1は活性化されず、ナトリウム輸送も起こらない。
[研究の経緯へ戻る]
発表論文
Diurnal regulation of SOS pathway and sodium excretion underlying salinity tolerance of Vigna marina (2024) Noda Y, Wang F, Chankaew S, Ariga H, Muto C, Iki Y, Ohashi H, Takahashi Y, Sakai H, Iseki K, Ogiso-Tanaka E, Suzui N, Yin YG, Miyoshi Y, Enomoto K, Kawachi N, Somta P, Furukawa J, Tomooka N, Naito K. Plant, Cell & Environment https://doi.org/10.1111/pce.15402
Multi-layered apoplastic barrier underlying the ability of Na+ exclusion in Vigna marina .(2024) Wang F, Tanoi, K, Yamauchi T, Naito, K. Plant and Cell Phisiology, https://doi.org/10.1093/pcp/pcaf005