日本に自生している野生植物で最も多い花色は白色と黄色で、それぞれ約3割を占め、次いで青・青紫・赤紫が約2割を占めます。赤や橙色の花色を持つ植物はほんのわずかで、ヤブツバキ(赤色)(図18)、コオニユリ(橙色)(図19)、マツモトセンノウ(橙色)(図20)など数えるほどしかありません。一方、花屋には様々な橙色の花が並んでいます。その多くは育種によって野生種にはないものが作られた結果です。



アントシアニンとカロテノイドによる橙色花色

<キク>
栽培ギク品種には白色や黄色の他に赤紫色や橙色の花色がありますが、野生のキク属には橙色のものはありません。アントシアニンにより赤紫の花色をしたキクとカロテノイドにより黄色の花色をしたキクを交雑し、両方の色素を持ち橙色をしたキクが作られたのです(図21、成果情報1)。

キク以外にもバラ(図22)、チューリップの橙色はアントシアニンとカロテノイドが合わさって発現しています。やはりキクと同じように橙色の花弁をもつ野生種は無く、交雑により両方の色素を持つように育種されました。
野生植物の中で、アントシアニンとカロテノイドの両方の色素を花弁に蓄積する植物は、ほとんどありません。花の色は昆虫や鳥などの花粉を運ぶ動物(ポリネーター)を呼び寄せるために進化したものですが、ポリネーターを呼ぶためにはどれか一種類の色素があれば十分で、複数の色素を蓄えるメリットが無いからだと考えられます。
フラボノイドによる橙色花色
<カーネーション>
カーネーションの黄色の花はカルコンにより、橙色の花はカルコンとアントシアニンの組み合わせにより発色しています。カルコンはアントシアニン生合成系の中間産物で、カルコン異性化酵素(CHI)の働きにより一旦無色のナリンゲニンに変換され、さらにフラバノン3-水酸化酵素(F3H)やジヒドロフラボノール還元酵素(DFR)、アントシアニジン合成酵素(ANS)の働きによりアントシアニンが合成されます(図106)。
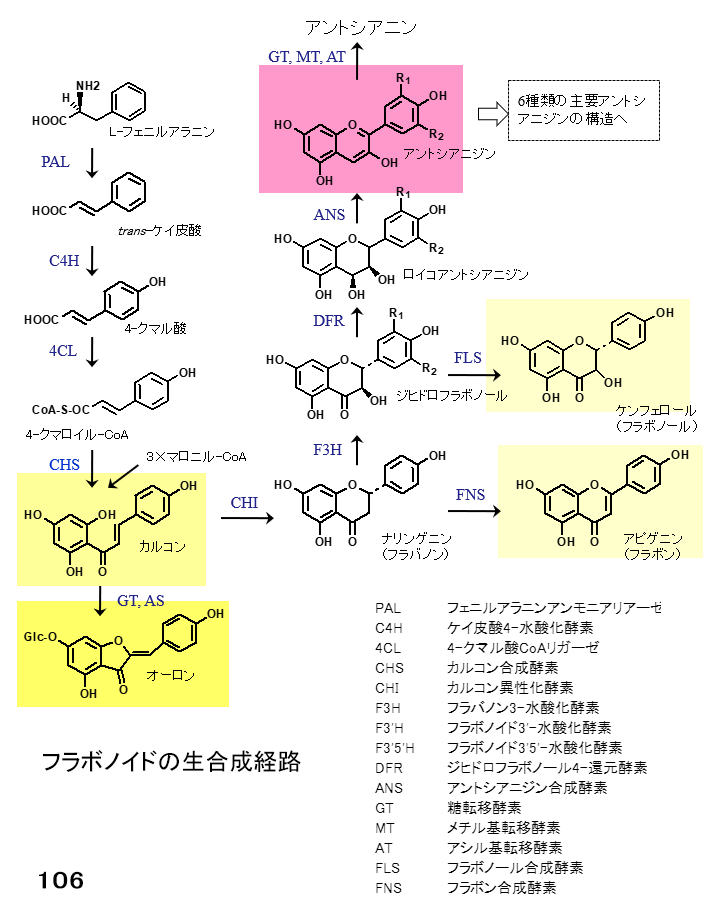{kind=link}
カーネーションの黄花や橙花にカルコンが貯まっているのは、CHI遺伝子の発現が低く抑えられているためです(図24)。この抑制は完全ではないので一部のカルコンはナリンゲニンに変換されます。黄色の花ではDFR遺伝子の発現も抑制されているため、ナリンゲニンからアントシアニンが合成されません。一方、橙色の花ではDFR酵素が正常に働いているので、わずかながらアントシアニンが合成され、カルコンの黄色とアントシアニンの赤色が混ざった橙色になります。
CHI遺伝子やDFR遺伝子の発現が抑制されるのは、これらの遺伝子にトランスポゾン※が挿入されているためです。カーネーションの野生種には、花弁にカルコンを貯めるものはありませんが、育種の過程でCHI遺伝子にトランスポゾンが入ったものが選抜されたと考えられます。
※トランスポゾン:「動く遺伝子」。遺伝子は普通決まった位置にありますが、トランスポゾンはいろいろなところに動くことが出来ます。色素を合成する酵素遺伝子の途中に、トランスポゾンが割り込んで入ると、その遺伝子が2つに分断され、酵素が働かなくなります(詳しくは「模様」の項を参照してください)。
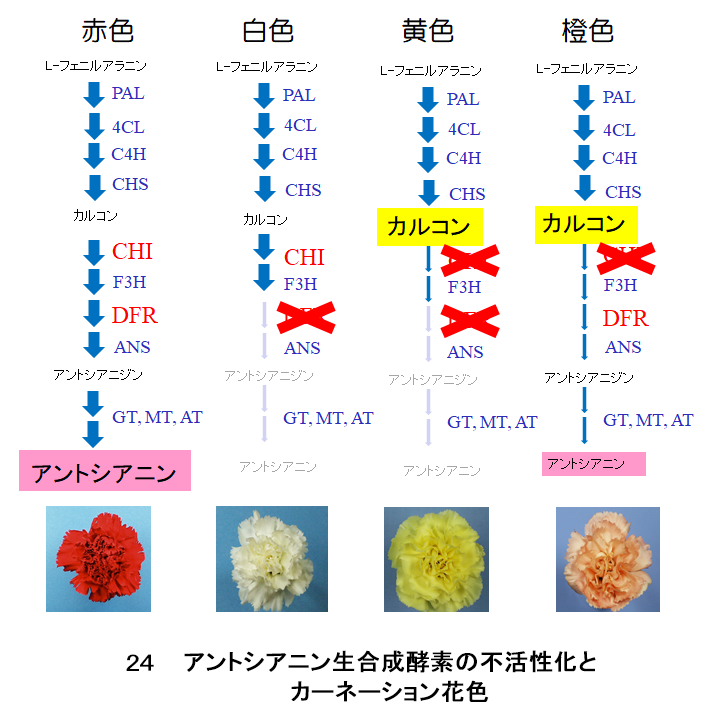
カロテノイドによる橙色花色
<マリーゴールド>
一種類の色素だけで、橙色を発色している植物もあります。マリーゴールドは、カロテノイドを多量に蓄積することで、橙色の花色を作っています。マリーゴールドの花弁に含まれるカロテノイドはルテインという黄色いカロテノイドですが、黄色と橙色の違いはルテインの量の違いで、橙色花弁は黄色花弁の何十倍ものルテインを含んでいます(図25、成果情報1)。黄色花弁と橙色花弁で生合成酵素遺伝子の発現を比較すると、橙色花弁の方が全体的に発現量が高く、特に生合成の鍵酵素と考えられているフィトエン合成酵素(PSY)の発現量が高いことが報告されています。


<キンセンカ>
キンセンカ(図26)には黄花品種と橙花品種があります。どちらもカロテノイドによって発色しています。キンセンカの場合、黄色花弁と橙色花弁に含まれるカロテノイドの量は同じですが、橙色花弁には黄色のカロテノイドの他に赤色のカロテノイド(リコペン類)が含まれています(成果情報1、成果情報2)。赤色のカロテノイドが貯まるのは、カロテノイド異性化酵素(CRTISO)の変異により、生合成が途中で止まってしまったためです(成果情報3)。
<ユリ>
ユリの場合も橙色花弁と黄色花弁(図27)ではカロテノイドの組成が異なります。黄色花弁はアンテラキサンチンやビオラキサンチンなど黄色のカロテノイドだけですが、橙色花弁にはカプサンチン(図27)と呼ばれる赤色のカロテノイドが含まれています。カプサンチンはトウガラシに含まれる赤色カロテノイドです。κ環と呼ばれる特殊な5員環構造を持っています。カプサンチンカプソルビン合成酵素の働きで、アンテラキサンチンから合成されます。花弁にカプサンチンを蓄積する例はユリ以外に報告されていません。

<カロテノイド分解産物による橙色>
カロテノイド分解産物のなかには、β-シトラウリンやクロセチンのように、橙色~赤色を発色するものがあります。このような分解産物を花弁の細胞に蓄積して、橙色を発色している植物があります。春咲きクロッカスやカリフォルニアポピー(図27b)、ヒオウギスイセン(図27c)の花弁の橙色は、クロセチンやその配糖体のクロシンにより発色しています。サフラン(クロッカスの雌しべ(写真27d)を乾燥させた香辛料)の赤色もクロセチンによるものです。



関連する農研機構成果情報(一般向け)
- 数種のキク科植物における黄色と橙色の花色発現に関わる色素組成 (2003)
- 橙色キンセンカの花色発現に関わるカロテノイド組成 (2004)
- キンセンカの花色はカロテノイド異性化酵素の活性によって決定される(2011)
専門家向け参考文献
- Kishimoto, S., Sumitomo, K., Yagi, M., Nakayama, M., Ohmiya, A. (2007) Three routes to orange petal color via carotenoid components in 9 Compositae species. J. Japan. Soc. Hort. Sci. 76: 250-257.
- Kishimoto, S., Ohmiya, A. (2009) Studies on carotenoids in the petals of Compositae plants. J. Japan. Soc. Hort. Sci.78: 263-272.
- Moehs, C. P., Tian, L., Osteryoung, K. W., DellaPenna, D. (2001) Analysis of carotenoid biosynthetic gene expression during marigold petal development. Plant Mol. Biol. 45: 281-293.
- Kishimoto, S., Maoka, T., Sumitomo, K., Ohmiya, A. (2005) Analysis of carotenoid composition in petals of calendula (Calendula officinalis L.). Biosci. Biotechnol. Biochem. 69: 2122-2128.
- Yamagishi, M., Kishimoto, S., Nakayama, M. (2009) Carotenoid composition and changes in expression of carotenoid biosynthetic genes in tepals of Asiatic hybrid lily. Plant Breeding, 129: 100-107.
- Itoh, Y., Higeta, D., Suzuki, A., Yoshida, H., Ozeki, Y. (2002) Excision of transposable elements from the chalcone isomerase and dihydroflavonol 4-reductase genes may contribute to the variegation of the yellow-flowered carnation (Dianthus caryophyllus). Plant Cell Physiol. 43: 578--585.
- 伊藤佳央ら (2004) 花の模様は遺伝子のしわざだった!化学、59: 40-43
- Rubio-Moraga, A., Ahrazem, O., Rambla, J. L., A. Granell, A., Gómez-Gómez, L. (2013) Crocins with high levels of sugar conjugation contribute to the yellow colours of early-spring flowering crocus tepals. PLoS ONE 8: e71946.